
Characterization of Polyphenols from Chenopodium botrys after Fractionation with Different Solvents and Study of Their In Vitro Biological Activity




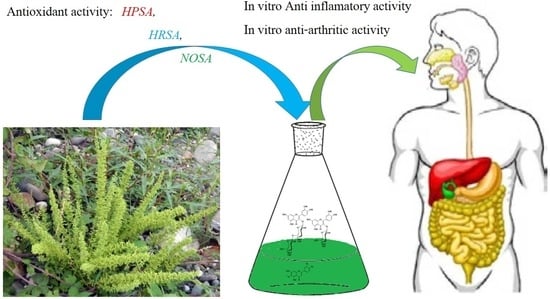
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Fractionation of Polyphenolic Complex with Different Solvents and Fraction Yields
2.2. Content of Total Phenols (TPC) and Tannins (TTC) in the Fractions
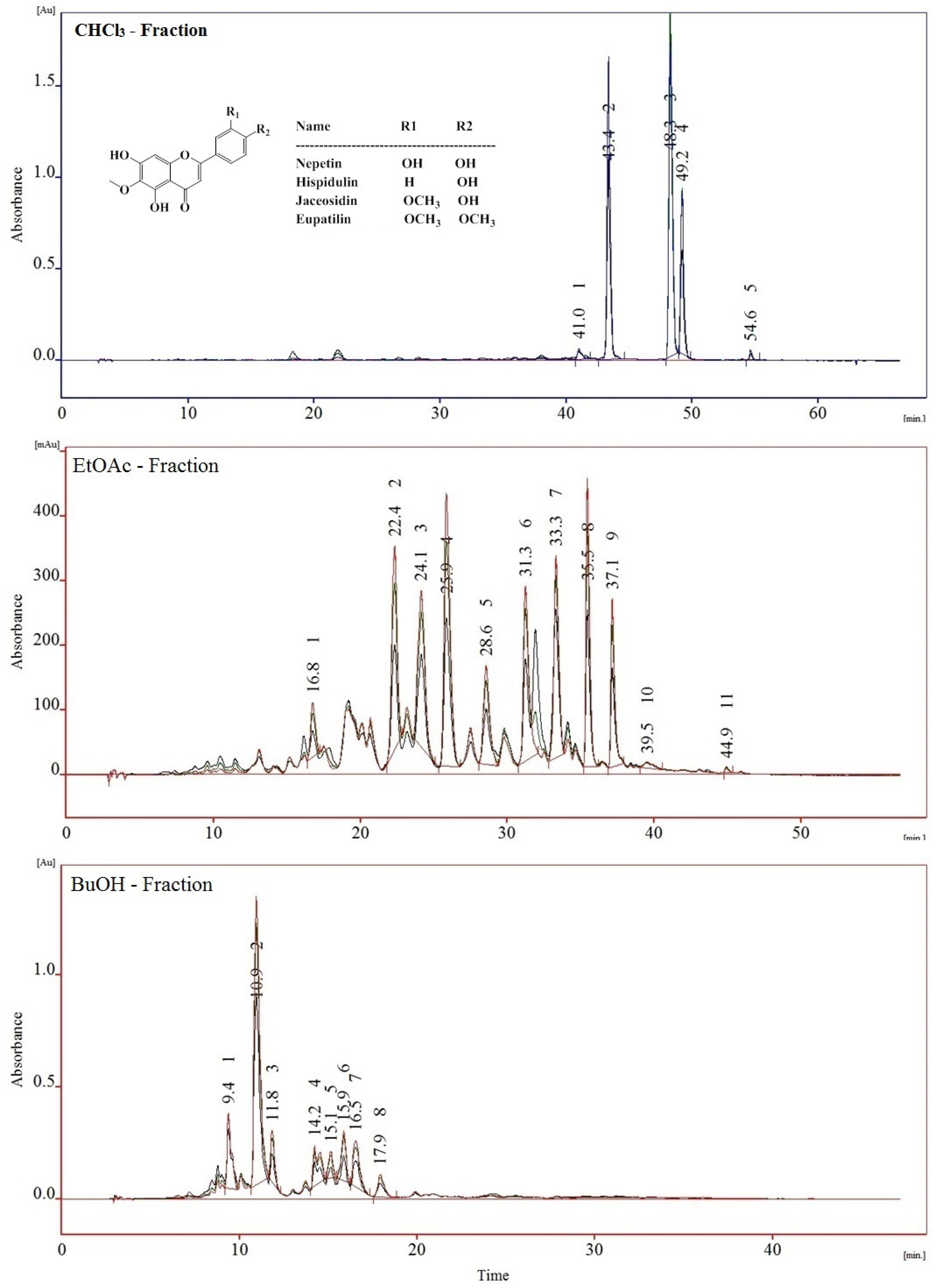
2.3. Content of 6-Methoxy Flavones and Quercetin Glycosides in the Fractions
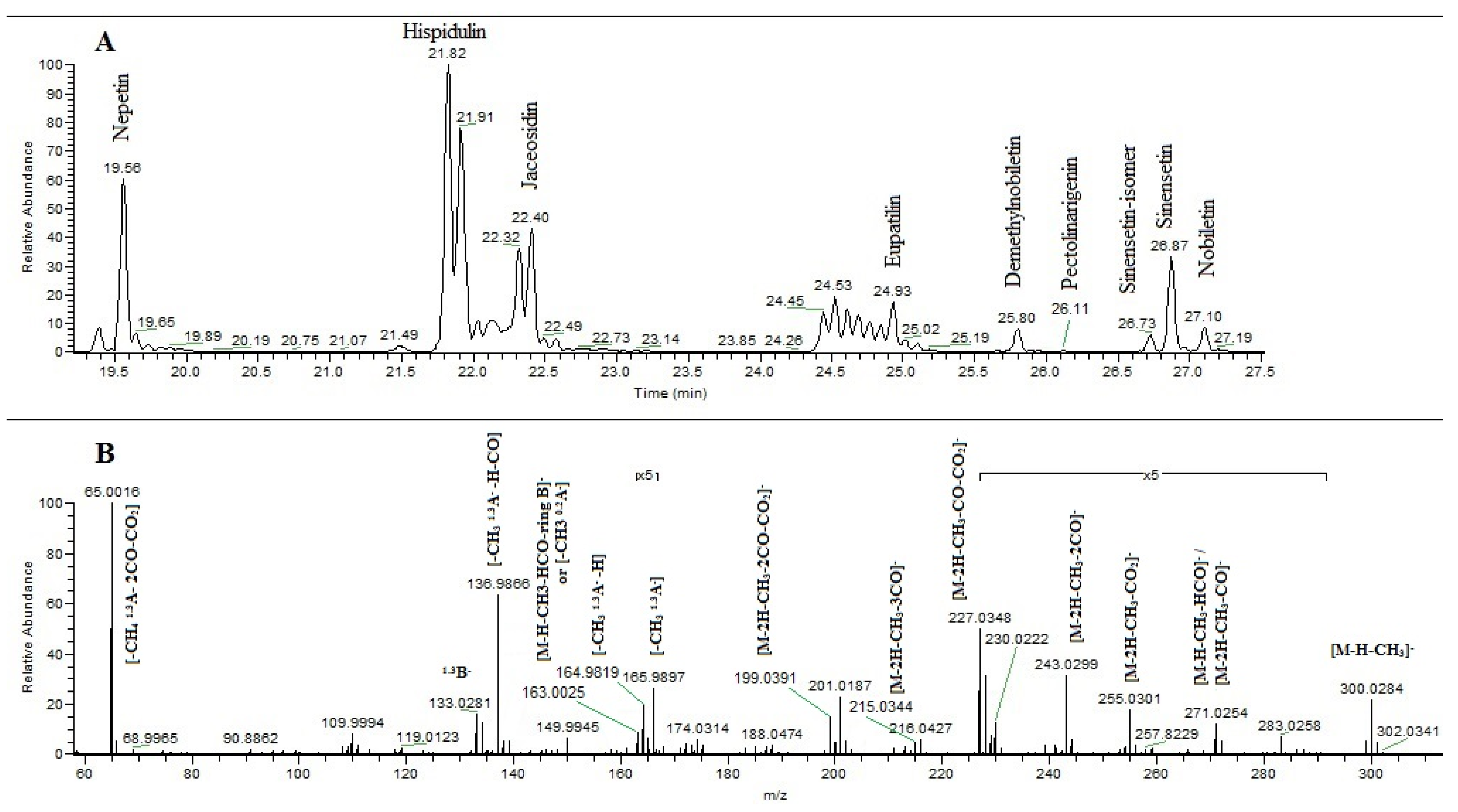
2.4. Identification of Polyphenolic Components by UHPLC-MS/MS
2.4.1. Flavonoids Identification
2.4.2. Identification of Glycosides
2.5. Hydrogen Peroxide Scavenging Activity
2.6. Hydroxyl Radical Scavenging Activity (HRSA)
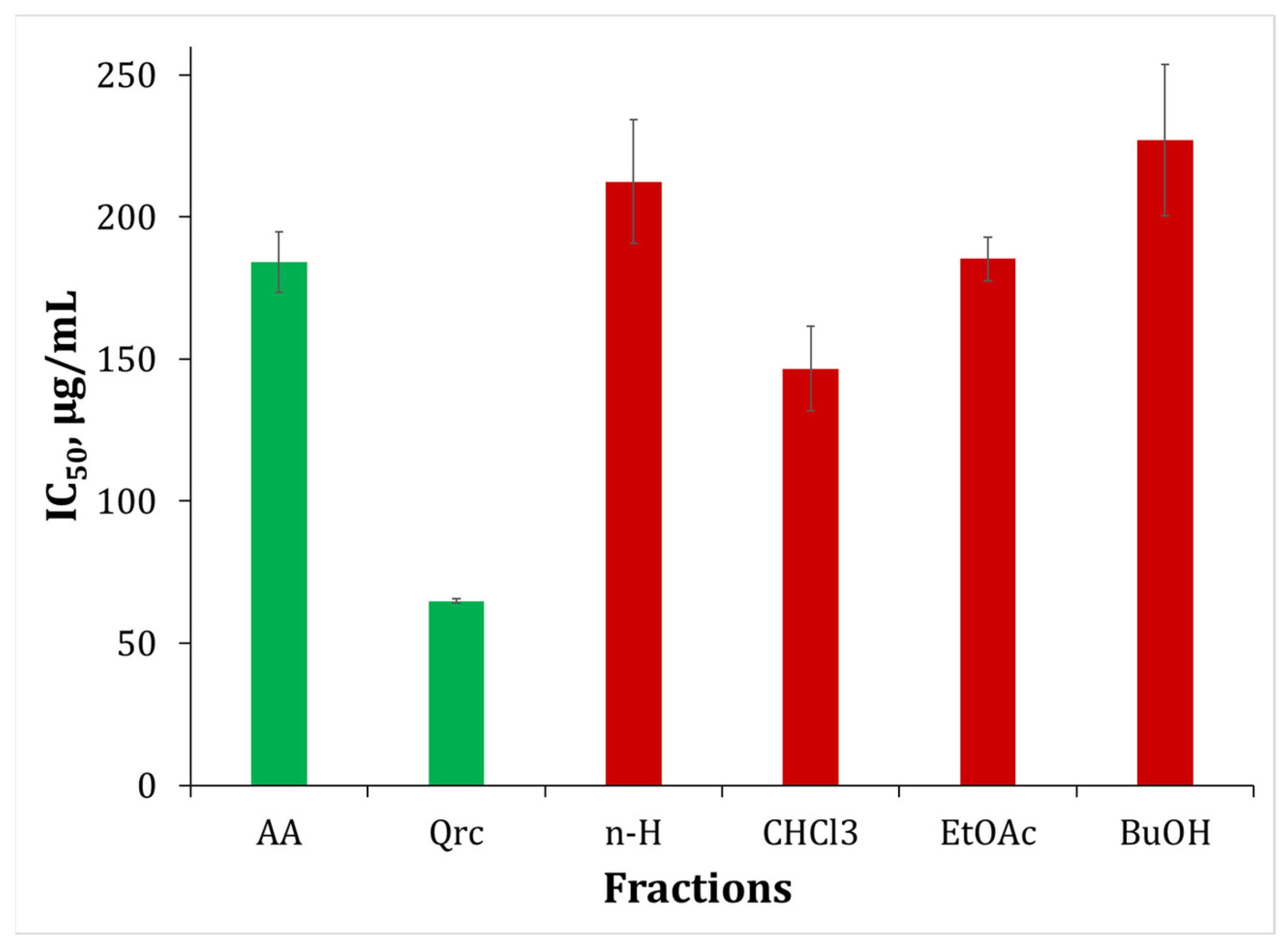
2.7. Nitric Oxide Scavenging Activity (NOSA)
2.8. Inhibition of Albumin Denaturation (IAD)
2.9. Antitryptic Activity (ATA)
2.10. Data Correlation
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Fractionation of Polyphenols with Solvents of Different Polarity
3.4. Determination of Total Phenolic Content (TPC)
3.5. Determination of Total Tannin Contain (TTC)
3.6. Analysis of Polyphenols by HPLC-PDA
3.7. Identification of Flavonoids by Orbitrap UHPLC-MS/MS
3.8. Methods for Investigation of Biological Activity
3.8.1. Hydrogen Peroxide Scavenging Activity (HPSA)
3.8.2. Hydroxyl Radical Scavenging Activity (HRSA)
3.8.3. Nitric Oxide Scavenging Activity (NOSA)
3.8.4. Inhibition of Albumin Denaturation (IAD)
3.8.5. Antitryptic Activity (ATA)
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Morteza-Semnani, K. A Review on Chenopodium botrys L.: Traditional uses, chemical composition and biological activities. Pharm. Biomed. Res. 2015, 1, 1–9. [Google Scholar] [CrossRef]
- Zargari, A. Medicinal Plants; Tehran University Publications: Tehran, Iran, 1993; Volume 4, pp. 218–219. [Google Scholar]
- Yadav, N.; Vasudeva, N.; Singh, S.; Sharma, S. Medicinal properties of genus Chenopodium Linn. NPR 2007, 6, 131–134. [Google Scholar]
- Maksimović, Z.; Dordević, S.; Mraović, M. Antimicrobial activity of Chenopodium botrys essential oil. Fitoterapia 2005, 76, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Bano, A.; Ahmad, M.; Hadda, T.; Saboor, A.; Sultana, S.; Zafar, M.; Khan, M.; Arshad, M.; Ashraf, M. Quantitative ethnomedicinal study of plants used in the skardu valley at high altitude of Karakoram-Himalayan range, Pakistan. J. Ethnobiol. 2014, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kletter, C.; Krichbaum, M. Tibetan Medicinal Plants; Medpharm Scientific Publishers: Guildford, UK, 2001; pp. 241–246. [Google Scholar]
- Dembitsky, V.; Shkrob, I.; Hanus, L. Ascaridole and related peroxides from the genus Chenopodium. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech. Repub. 2008, 152, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari-Karchegani, N.; Amjad, L.; Ranjbar, M. Comparative Analysis of Chemical Composition of three Ecotypes of Chenopodium. AEB 2014, 8, 43–47. [Google Scholar]
- Morteza-Semnani, K.; Babanezhad, E. Essential Oil Composition of Chenopodium botrys L. from Iran. J. Essent. Oil-Bear. Plants 2007, 10, 314–317. [Google Scholar] [CrossRef]
- Tzakou, O.; Pizzimenti, A.; Pizzimenti, P.; Sdrafkakis, V.; Galati, M. Composition and antimicrobial activity of Chenopodium botrys L. essential oil from Greece. J. Essent. Oil Res. 2007, 19, 292–294. [Google Scholar] [CrossRef]
- El-Sayed, A.; Al-Yahya, M.; Hassan, M. Chemical Composition and Antimicrobial Activity of the Essential Oil of Chenopodium botrys Growing in Saudi Arabia. Pharm. Biol. 1989, 27, 185–188. [Google Scholar] [CrossRef]
- Mahboubi, M.; Bidgoli, F.; Farzin, N. Chemical Composition and antimicrobial activity of Chenopodium botrys L. essential oil. J. Essent. Oil-Bear. Plants 2011, 14, 498–503. [Google Scholar] [CrossRef]
- Feizbakhsh, A.; Sedaghat, S.; Tehrani, M.; Rustaiyan, A.; Masoudi, S. Chemical Composition of the Essential Oils of Chenopodium botrys L. from Two Different Locations in Iran. J. Essent. Oil Res. 2003, 15, 193–194. [Google Scholar] [CrossRef]
- Bedrossian, A.; Beauchamp, P.; Bernichi, B.; Dev, V.; Kitaw, K.; Rechtshaffen, H.; Bottini, A.; Hope, H. Analysis of North American Chenopodium botrys essential oil isolation and structure of two new sesquiterpene alcohols. J. Essent. Oil Res. 2001, 13, 393–400. [Google Scholar] [CrossRef]
- Bojilov, D.; Dagnon, S.; Ivanov, I. Constituent composition of Chenopodium botrys essential oil. Bulg. Chem. Commun. 2017, 49, 124–129. Available online: http://www.bcc.bas.bg/BCC_Volumes/Volume_49_Special_G_2017/BCC-49-G-Bojilov-124-129.pdf (accessed on 19 April 2023).
- Bahrman, N.; Jay, M.; Gorenflot, R. Contribution to the chemosystematic knowledge of some species of the genus Chenopodium L. Lett. Bot. 1985, 2, 107–113. [Google Scholar]
- de Pascual, T.J.; González, M.; Vicente, S.; Bellido, I. Flavonoids from Chenopodium botrys. Planta Med. 1981, 41, 389–391. [Google Scholar] [CrossRef]
- Rustembekowa, G.; Goryaev, M.; Nezhinskaya, G. Flavonoids of Chenopodium botrys. Chem. Nat. Compd. 1974, 10, 406. [Google Scholar] [CrossRef]
- Bojilov, D.; Dagnon, S.; Ivanov, I. New insight into the flavonoid composition of Chenopodium botrys. Phytochem. Lett. 2017, 20, 316–321. [Google Scholar] [CrossRef]
- Arisawa, M.; Minabe, N.; Saeki, R.; Takakuwa, T.; Nakaoki, T. Studies on unutilized resources. V. The components of the flavonoids in Chenopodium genus plants. 1. Flavonoids of Chenopodium ambrosioides L. Yakugaku Zasshi 1971, 91, 522–524. [Google Scholar] [CrossRef] [Green Version]
- Bylka, W.; Kowalewski, Z. Flavonoids in Chenopodium album L. and Chenopodium opulifolium L. Herba Pol. 1997, 43, 208–213. [Google Scholar]
- Chludil, H.; Corbino, G.; Leicach, S. Soil Quality Effects on Chenopodium album Flavonoid Content and Antioxidant Potential. J. Agric. Food Chem. 2008, 56, 5050–5056. [Google Scholar] [CrossRef]
- De Smone, A.; Dini, A.; Saturnino, P.; Schettino, O. Two flavonolglycosides from Chenopodium quinoa. Phytochemistry 1990, 29, 3690–3692. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Tenore, G.; Dini, A. Phenolic constituents of Kancolla seeds. Food Chem. 2004, 84, 163–168. [Google Scholar] [CrossRef]
- El-Sayed, N.; Awaad, A.; Hifnawy, M.; Mabry, T. A Flavonol Triglucoside from Chenopodium murale. Phytochemistry 1999, 51, 591–593. [Google Scholar] [CrossRef]
- Ghareeb, M.; Saad, A.; Abdou, A.; Refahy, L.; Ahmed, W. A new kaempferol glycoside with antioxidant activity from Chenopodium ambrosioides growing in Egypt. Orient 2016, 32, 3053–3061. [Google Scholar] [CrossRef] [Green Version]
- Gohar, A.; Elmazar, M. Isolation of hypotensive flavonoids from Chenopodium species growing in Egypt. Phytother. Res. 1997, 11, 564–567. [Google Scholar] [CrossRef]
- Gohar, A.; Maatooq, G.; Niwa, M. Two flavonoid glycosides from Chenopodium murale. Phytochemistry 2000, 53, 299–303. [Google Scholar] [CrossRef]
- Hammoda, H.; Harraz, F.; El Ghazouly, M.; Radwan, M.; Elsohly, M.; Wanas, A.; Bassam, S. Two new flavone glycosides from Chenopodium ambrosioides growing wildly in Egypt. Rec. Nat. Prod. 2015, 9, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Fujita, T.; Ishii, T.; Ueno, N. Antioxidative properties and flavonoid composition of Chenopodium quinoa seeds cultivated in Japan. Food Chem. 2010, 119, 1300–1306. [Google Scholar] [CrossRef]
- Ibrahim, L.; Kawashty, S.; Baiuomy, A.; Shabana, M.; El-Eraky, W.; El-Negoumy, S. A comparative study of the flavonoids and some biological activities of two Chenopodium species. Chem. Nat. Compd. 2007, 43, 24–28. [Google Scholar] [CrossRef]
- Jain, N.; Alam, M.; Kamil, M.; Ilyas, M.; Niwa, M.; Sakae, A. Two flavonol glycosides from Chenopodium ambrosioides. Phytochemistry 1990, 29, 3988–3991. [Google Scholar] [CrossRef]
- Kokanova-Nedialkova, Z.; Nedialkov, P.; Nikolov, S. The Genus Chenopodium: Phytochemistry, Ethnopharmacology and Pharmacology. Phcog. Rev. 2009, 3, 280–306. [Google Scholar]
- Rahiminejad, M.; Gornall, R. Flavonoid evidence for allopolyploidy in the Chenopodium album aggregate (Amaranthaceae). Plant Syst. Evol. 2004, 246, 77–87. [Google Scholar] [CrossRef]
- Rastrelli, L.; Saturnino, P.; Schettino, O.; Dini, A. Studies on the Constituents of Chenopodium pallidicaule (Canihua) Seeds. Isolation and Characterization of Two New Flavonol Glycosides. J. Agric. Food Chem. 1995, 43, 2020–2024. [Google Scholar] [CrossRef]
- Zhu, N.; Sheng, S.; Li, D.; Lavoie, E.; Karwe, M.; Rosen, R.; Ho, C.-T. Antioxidative flavonoid glycosides from quinoa seeds (Chenopodium quinoa wild). J. Food Lipids 2001, 8, 37–44. [Google Scholar] [CrossRef]
- Chalana, A.; Kumar Rai, R.; Karri, R.; Kumar Jha, K.; Kumar, B.; Roy, G. Interplay of the intermolecular and intramolecular interactions in stabilizing the thione-based copper(I) complexes and their significance in protecting the biomolecules against metal-mediated oxidative damage. Polyhedron 2022, 215, 115647. [Google Scholar] [CrossRef]
- Galano, A.; Macías-Ruvalcaba, N.; Campos, O.; Pedraza-Chaverri, J. Mechanism of the OH radical scavenging activity of nordihydroguaiaretic acid: A combined theoretical and experimental study. J. Phys. Chem. B 2010, 114, 6625–6635. [Google Scholar] [CrossRef]
- Halliwell, B. Gutterdge Free Radicals in Biology and Medicine; Clarendon Press: Oxford, UK, 1985; p. 346. [Google Scholar]
- Russo, D. Flavonoids and the Structure-Antioxidant Activity Relationship. J. Pharmacogn. Nat. Prod. 2018, 4, 30–31. [Google Scholar] [CrossRef]
- Xiao, J. Dietary flavonoid aglycones and their glycosides: Which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef]
- Bektaşoǧlu, B.; Esin Çelik, S.; Özyürek, M.; Güçlü, K.; Apak, R. Novel hydroxyl radical scavenging antioxidant activity assay for water-soluble antioxidants using a modified CUPRAC method. Biochem. Biophys. Res. Commun. 2006, 345, 1194–1200. [Google Scholar] [CrossRef]
- Treml, J.; Šmejkal, K. Flavonoids as Potent Scavengers of Hydroxyl Radicals. Compr. Rev. Food Sci. Food Saf. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Jagetia, G.; Baliga, M. The evaluation of nitric oxide scavenging activity of certain Indian medicinal plants in vitro: A preliminary study. J. Med. Food 2004, 7, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Lala, P. Significance of nitric oxide in carcinogenesis, tumor progression and cancer therapy. Cancer Metastasis Rev. 1998, 17, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Morikawa, T.; Ando, S.; Toguchida, I.; Yoshikawa, M. Structural Requirements of Flavonoids for Nitric Oxide Production Inhibitory Activity and Mechanism of Action. Bioorg. Med. Chem. 2003, 11, 1995–2000. [Google Scholar] [CrossRef] [PubMed]
- Osman, N.; Sidik, N.; Awal, A.; Adam, N.; Rezali, N. In vitro xanthine oxidase and albumin denaturation inhibition assay of Barringtonia racemosa L. and total phenolic content analysis for potential anti-inflammatory use in gouty arthritis. J. Intercult. 2016, 5, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Opie, E. On the relation of necrosis and inflammation to denaturation of proteins. J. Exp. Med. 1962, 115, 597–608. [Google Scholar] [CrossRef]
- Oyedapo, O.; Famurewa, A. Antiprotease and membrane stabilizing activities of extracts of fagara zanthoxyloides, olax subscorpioides and tetrapleura tetraptera. Int. J. Pharmacogn. 1995, 33, 65–69. [Google Scholar] [CrossRef]
- Xue, G.; Gong, L.; Yuan, C.; Xu, M.; Wang, X.; Jiang, L.; Huang, M. A structural mechanism of flavonoids in inhibiting serine proteases. Food Funct. 2017, 8, 2437–2443. [Google Scholar] [CrossRef]
- Singleton, V.; Orthofer, R.; Lamuela-Ravent, R. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Bojilov, D.; Manolov, S.; Bazadzhiev, B.; Stremski, J.; Ivanov, I. Antioxidant activity in two species common beans (Phaseolus vulgaris L.) from village of Smilyan, Bulgaria. J. Int. Sci. Publ. Agric. Food 2018, 6, 314–324. Available online: https://www.scientific-publications.net/en/article/1001751/ (accessed on 21 April 2023).
- Haile, M.; Kang, W. Antioxidant Activity, Total Polyphenol, Flavonoid and Tannin Contents of Fermented Green Coffee Beans with Selected Yeasts. Fermentation 2019, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Manolov, S.; Ivanov, I.; Bojilov, D. Synthesis of New 1,2,3,4-Tetrahydroquinoline Hybrid of Ibuprofen and Its Biological Evaluation. Molbank 2022, 2022, M1350. [Google Scholar] [CrossRef]
- Guo, T.; Wei, L.; Sun, J.; Hou, C.; Fan, L. Antioxidant activities of extract and fractions from Tuber indicum Cooke & Massee. Food Chem. 2011, 127, 1634–1640. [Google Scholar] [CrossRef]
- Marcocci, L.; Maguire, J.; Droylefaix, M.; Packer, L. The Nitric Oxide-Scavenging Properties of Ginkgo Biloba Extract EGb 761. Biochem. Biophys. Res. Commun. 1994, 201, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Manolov, S.; Ivanov, I.; Bojilov, D. Microwave-assisted synthesis of 1,2,3,4-tetrahydroisoquinoline sulfonamide derivatives and their biological evaluation. J. Serb. Chem. Soc. 2021, 86, 139–151. [Google Scholar] [CrossRef]
- Nijveldt, R.; van Nood, E.; van Hoorn, D.; Boelens, P.; van Norren, K.; van Leeuwen, P. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [Green Version]
- Shoskes, D. Effect of bioflavonoids quercetin and curcumin on ischemic renal injury: A New Class of Renoprotective Agents: 1. Transplantation 1998, 66, 147–152. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions | Yield | TPC | TTC | Qrc-Glc | 6-MFs |
|---|---|---|---|---|---|
| % | mgGAE/g Extr | mgTAE/g Extr | mg/g Extr | mg/g Extr | |
| n-H | 33.02 ± 0.71 | 18.41 ± 1.76 | 10.40 ± 0.79 | - | - |
| CHCl3 | 12.30 ± 0.71 | 48.92 ± 2.26 | 36.97 ± 3.05 | - | 355.47 ± 7.14 |
| EtOAc | 10.74 ± 0.94 | 159.84 ± 2.59 | 133.08 ± 14.56 | 168.82 ± 3.73 | - |
| BuOH | 24.93 ± 1.17 | 50.05 ± 2.06 | 33.85 ± 1.41 | 67.21 ± 1.56 | - |
| Nepetin | Hispidulin | Jaceosidin | Eupatilin | Pectolinarigenin | |
|---|---|---|---|---|---|
| [M–H]− | 315.0515 [−3.17] a | 299.0573 [−5.68] | 329.0671 [−3.04] | 343.0831 [−3.79] | 313.0732 [4.47] |
| [M–H–CH3]− | 300.0275 (22.4) b | 284.0331 (20.4) | 314.0437 (2.6) | 328.0598 (1.2) | - |
| [M–H–CH4]− | - | - | 313.0363 (3.1) | - | - |
| [M–H–2CH3]− | - | - | 299.0207 (16.4) | 313.0364 (37.6) | 283.0254 (80.24) |
| [M–H–CH3–HCO]−/[M–2H–CH3–CO]− | 271.0243 (2.8) | 255.0301 (4) | 285.0412 (2) | - | - |
| [M–H–3CH3]− | - | - | - | 298.0127 (100) | - |
| [M–H–2CH3–CO]− | - | - | 271.0251 (100) | 285.0413 (14.3) | 255.0301 (77.31) |
| [M–H–3CH3–CO]− | - | - | - | 270.0174 (65.7) | - |
| [M–2H–CH3–CO2]− | 255.0299 (2.8) | - | - | - | - |
| [M–H–3CH3–CO2]− | - | - | - | 254.0223 (3.6) | - |
| [M–H–2CH3–2CO]− | - | - | 243.0299 (6.8) | - | 227.0347 (17.32) |
| [M–H–3CH3–2CO]− | - | - | 242.0221 (27.7) | - | |
| [M–2H–CH3–2CO]− | 243.0301 (5.3) | 227.0347 (14.6) | - | - | - |
| [M–H–2CH3–CO–CO2]− | - | - | 227.0347 (5.7) | - | 211.0401 (12) |
| [M–H–3CH3–CO–CO2]− | - | - | 226.0269 (8.9) | - | |
| [M–H–CH3–C2H2O–CO]− | - | 214.0267 (1) | - | - | - |
| [M–2H–CH3–CO–CO2]− | 227.0348 (9.7) | 211.0391 (14.3) | - | - | - |
| [M–H–2CH3–3CO]− | – | - | 215.0345 (4.6) | - | 199.0392 (3.5) |
| [M–H–3CH3–3CO]− | - | - | 214.0267 (11.8) | - | |
| [M–2H–CH3–3CO]− | 215.0343 (4.6) | 199.0394 (7.7) | - | - | - |
| [M–H–2CH3–2CO–CO2]− | - | - | 199.0395 (16.7) | - | 183.0442 (33) |
| [M–H–3CH3–2CO–CO2]− | - | - | 198.0315 (13.4) | - | |
| [M–2H–CH3–2CO–CO2]− | 199.0392 (15.7) | 183.0439 (11) | - | - | - |
| [M–H–2CH3–4CO]− | - | - | 187.0391 (2.3) | - | - |
| [M–H–3CH3–4CO]− | - | - | 186.0317 (2.5) | - | |
| [M–H–2CH3–3CO–CO2]− | - | - | 171.0441 (5.4) | - | - |
| [M–H–3CH3–3CO–CO2]− | - | - | 170.0363 (5.1) | - | |
| CH3 1.3A− | 165.9896 (3.5) | 165.9894 (2.7) | 165.9899 (1.7) | 165.9900 (3.7) | 165.9894 (4) |
| [CH3 1.3A–H]− | 164.9818 (2.2) | - | 164.9818 (2.2) | 164.9818 (11.2) | 164.9817 (12.53) |
| [CH3 0.2A−]/[M–H–CH3–HCO–ring B]− | 163.0026 (5) | - | 163.0026 (2.6) | 163.0025 (34) | 163.0025 (72) |
| [CH3 1.3A–CO]− | 136.9866 (63) | 136.9866 (84) | 136.9866 (12) | 136.9866 (16.8) | 136.9866 (20.02) |
| [CH30.2A−–CO]/[M–H–CH3–HCO–ring B–CO]− | - | - | 135.0037 (2) | 135.0037 (28) | 135.0037 (40) |
| 1.3B− | 133.0290 (17.8) | 117.0330 (8.2) | 147.0439 (3.2) | - | - |
| CH3 1.3B− + H | - | - | 133.0281 (9.8) | - | - |
| CH3 1.3B− | - | - | 132.0202 (1) | 147.0438 (30.5) | 117.0331 (67.66) |
| 2CH3 1.3B− | - | - | - | 132.0202 (46) | - |
| [CH3 1.3A–2CO–CO2]− | 65.0016 (100) | 65.0016 (100) | 65.0016 (25) | 65.0015 (16) | 65.0016 (28.18) |
| Fraction | Peak № | Name | UHPLC-MS | [M–H]− | MS/MS |
|---|---|---|---|---|---|
| tR | (Δm, ppm) | ||||
| BuOH fraction | 1 | Quercetine 3-O-(2″-O-apiofuranosyl-6″-O-glucopyranosyl)- β-D-glucopyranoside | 12.11 | 757.1839 (0.79) | 625, 463, 300, 271, 255, 243, 179, 151, 112, 121, 107 |
| 2 | Quercetine 3-O-(2″,6″-di-O-α-L-rhamnopyranosyl)-β-D- galactopyranoside (acyl: malonyl) | 12.36 | 841.2052 (0.95) | 463, 300, 271, 255, 243, 179, 151, 121, 112, 107 | |
| 3 | Quercetine 3-O-(2″,6″-di-O-α-L-rhamnopyranosyl)- β-D-glucopyranoside (acyl: malonyl) | 12.70 | 841.2052 (0.95) | 463, 300, 271, 255, 243, 179, 151, 121, 112, 107 | |
| 4 | Quercetine 3-O-α-L-rhamnopyranosyl-α-L-rhamnopyranosyl- β-D-malonylglucopyranoside | 13.69 | 841.2052 (0.95) | 463, 300, 271, 255, 243, 179, 151, 121, 112, 107 | |
| 5 | Quercetine 3-O-α-L-rhamnopyranosyl-α-L-rhamnopyranosyl- β-D-malonylglucopyranoside (acyl) | 13.77 | 883.2162 (1.02) | 841, 737, 463, 300, 271, 255, 243, 179, 151, 121, 107 | |
| EtOAc fraction | 6 | Rutin | 12.14 | 609.1467 (0.98) | 300, 271, 255, 243, 179, 151, 107 |
| 7 | Quercetin-O-pentoside-hexoside | 12.22 | 595.1310 (0.84) | 300, 271, 255, 243, 179, 151, 107 | |
| 8 | Quercetin-O-(acyl)-hexoside-hexoside | 12.31 | 667.1524 (1.20) | 625, 463, 300, 271, 255, 243, 179, 151, 107 | |
| 9 | Quercetin-O-(acyl)-hexoside-(acyl)-hexoside | 12.48 | 709.1622 (0.00) | 667, 639, 505, 463, 300, 271, 255, 243, 179, 151, 107 | |
| 10 | Isorhamnetin 3-O-glucopyranoside-7-O-rhamnopyranoside | 12.56 | 623.1628 (−1.60) | 461, 314, 300, 285, 271, 255, 243, 227, 165, 151, 137, 125, 107 | |
| 11 | Quercetin 3-O-galactopyranoside | 12.81 | 463.0881 (−0.22) | 301, 300, 271, 255, 243, 179, 151, 107 | |
| 12 | Quercetin 3-O-glucopyranoside | 12.90 | 463.0881 (−0.22) | 301, 300, 271, 255, 243, 179, 151, 107 | |
| 13 | Isorhamnetin 3-O-glucopyranoside-rhamnopyranoside | 13.92 | 623.1628 (−1.60) | 477, 315, 300, 285, 271, 255, 243, 227, 165, 151, 137, 125, 107 | |
| 14 | Jaceosidin 7-O-glucopyranosid | 13.94 | 491.1197 (0.41) | 328, 313, 300, 285, 269, 241, 213, 164, 147, 133 | |
| 15 | Quercetin-O-(acyl)-hexoside-(acyl)-hexoside | 14.01 | 709.1613 (−1.27) | 667, 463, 300, 271, 255, 243, 179, 151, 107 | |
| 16 | Isorhamnetin 3-O-glucopyranoside | 14.26 | 477.1035 (−0.63) | 315, 314, 299, 285, 271, 257, 243, 227, 215, 199, 151, 125, 107 | |
| 17 | Hispidulin 7-O-glucopyranoside | 14.6 | 461.1084 (−1.08) | 297, 283, 269, 255, 227, 183, 164, 163, 136, 135, 117 | |
| 18 | Kaempferol-O-(acyl)-hexoside-(acyl)-hexoside | 14.85 | 693.1680 (1.15) | 489, 447, 285, 255, 227, 213, 151, 125, 107 | |
| 19 | Jaceosidin 4′-O-glucopyranosid | 14.94 | 491.1191 (−0.81) | 328, 313, 298, 285, 270, 161, 164, 163, 136, 135, 132 | |
| 20 | Quercetin-glycoside—unidentified | 15.03 | 883.2127 | 300, 271, 255, 243, 179, 151, 107 | |
| 21 | Quercetin-glycoside—unidentified | 15.71 | 853.2035 | 300, 271, 255, 243, 227, 151, 107 | |
| 22 | Kaempferol-glycoside—unidentified | 15.88 | 735.1791 | 443, 285, 284, 255, 227, 213, 151, 121, 125, 107 | |
| 23 | Quercetin-glycoside—unidentified | 16.22 | 751.1723 | 300, 271, 255, 243, 227, 179, 151, 121, 107 | |
| 24 | Quercetin-glycoside—unidentified | 16.40 | 751.1740 | 300, 271, 255, 243, 227, 179, 151, 121, 107 | |
| 25 | Quercetin-glycoside—unidentified | 16.82 | 925.2233 | 300, 271, 255, 243, 179, 151, 107 | |
| 26 | Quercetin-glycoside—unidentified | 17.67 | 793.1818 | 300, 271, 255, 243, 179, 151, 121, 107 |
| Fractions | HPSA | HRSA | NOSA | IAD | ATA |
|---|---|---|---|---|---|
| IC50 ± SD, μg/mL | |||||
| n-H | 511.49 ± 33.33 | 149.57 ± 2.02 | 212.45 ± 21.76 | 945.06 ± 86.88 | 560.99 ± 15.24 |
| CHCl3 | 417.03 ± 34.07 | 139.32 ± 8.85 | 146.59 ± 14.94 | 840.38 ± 60.03 | 202.44 ± 23.36 |
| EtOAc | 39.18 ± 3.26 | 105.03 ± 3.52 | 185.25 ± 7.63 | 461.41 ± 43.84 | 116.23 ± 11.59 |
| BuOH | 978.07 ± 136.03 | 143.76 ± 3.46 | 227.12 ± 26.61 | 848.38 ± 68.93 | 145.56 ± 4.73 |
| Standard | |||||
| AA | 24.84 ± 0.35 | 129.84 ± 4.52 | 184.11 ± 10.78 | - | - |
| Qrc | 69.25 ± 1.82 | 149.18 ± 9.27 | 64.82 ± 0.74 | - | - |
| Ibu | - | - | - | 81.50 ± 4.95 | 259.82 ± 9.14 |
| Ket | - | - | - | 126.58 ± 5.00 | 720.57 ± 19.78 |
| TTC | Qrc-Glc | 6-MFs | HPSA | HRSA | NOSA | IAD | ATA | |
|---|---|---|---|---|---|---|---|---|
| TPC | 0.9993 | 0.9412 | −0.2177 | −0.6916 | −0.9900 | −0.2133 | −0.9996 | −0.6511 |
| TTC | 1 | 0.9306 | −0.2038 | −0.7183 | −0.9941 | −0.2356 | −1.0000 | −0.6312 |
| Qrc-Glc | 1 | −0.4931 | −0.4596 | −0.8853 | 0.1204 | −0.9336 | −0.6299 | |
| 6-MFs | 1 | −0.1199 | 0.1139 | −0.8716 | 0.2078 | −0.1741 | ||
| HPSA | 1 | 0.7773 | 0.5843 | 0.7115 | 0.0819 | |||
| HRSA | 1 | 0.3343 | 0.9931 | 0.6126 | ||||
| NOSA | 1 | 0.2296 | 0.2618 | |||||
| IAD | 1 | 0.6362 | ||||||
| ATA | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bojilov, D.; Manolov, S.; Nacheva, A.; Dagnon, S.; Ivanov, I. Characterization of Polyphenols from Chenopodium botrys after Fractionation with Different Solvents and Study of Their In Vitro Biological Activity. Molecules 2023, 28, 4816. https://doi.org/10.3390/molecules28124816
Bojilov D, Manolov S, Nacheva A, Dagnon S, Ivanov I. Characterization of Polyphenols from Chenopodium botrys after Fractionation with Different Solvents and Study of Their In Vitro Biological Activity. Molecules. 2023; 28(12):4816. https://doi.org/10.3390/molecules28124816
Chicago/Turabian StyleBojilov, Dimitar, Stanimir Manolov, Angelika Nacheva, Soleya Dagnon, and Iliyan Ivanov. 2023. "Characterization of Polyphenols from Chenopodium botrys after Fractionation with Different Solvents and Study of Their In Vitro Biological Activity" Molecules 28, no. 12: 4816. https://doi.org/10.3390/molecules28124816