Climate Change and Alpine Screes: No Future for Glacial Relict Papaver occidentale (Papaveraceae) in Western Prealps

 , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
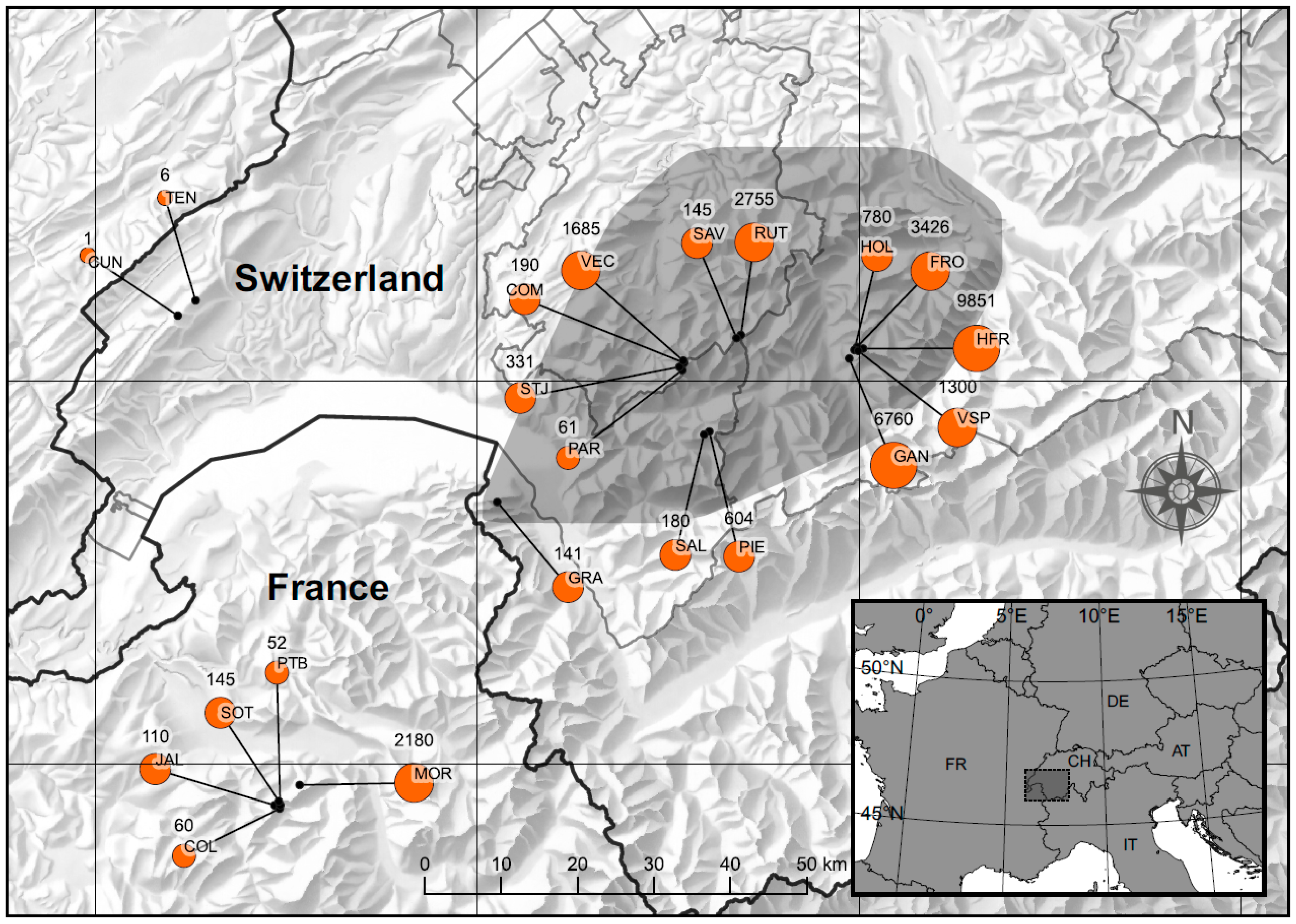
2.1. Study Area and Data Collection
2.2. Environmental Variables
2.3. Model Selection and Calibration
2.4. Model Validation
3. Results
3.1. Distribution and Population Size of Papaver occidentale
3.2. Species Distribution Modelling
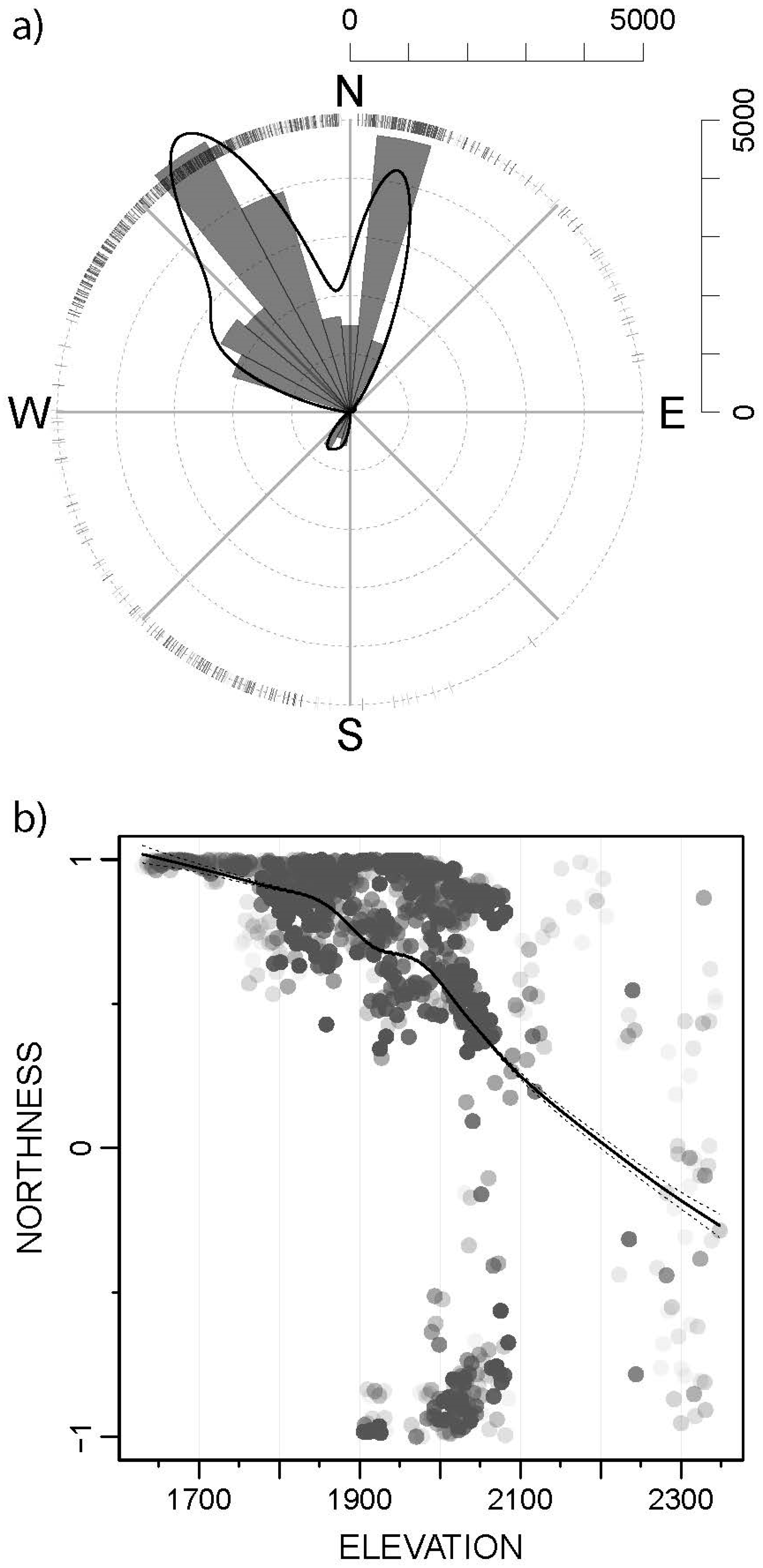
3.3. Predicted Habitat Suitability and Ecological Preferences
4. Discussion
4.1. Current Geographic Range
4.2. No Future for P. occidentale?
4.3. Conservation Implications
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiménez-Alfaro, B.; García-Calvo, L.; García, P.; Luis Acebes, J. Anticipating extinctions of glacial relict populations in mountain refugia. Biol. Conserv. 2016, 201, 243–251. [Google Scholar] [CrossRef]
- Arrigo, N.; Bétrisey, S.; Graf, L.; Bilat, J.; Gerber, E.; Kozlowski, G. Hybridization as a threat in climate relict Nuphar pumila (Nymphaeaceae). Biodiv. Conserv. 2016, 25, 1863–1877. [Google Scholar] [CrossRef] [Green Version]
- Zecca, G.; Casazza, G.; Piscopo, S.; Minuto, L.; Grassi, F. Are the responses of plant species to Quaternary climatic changes idiosyncratic? A demographic perspective from Western Alps. Plant Ecol. Divers. 2017, 10, 273–281. [Google Scholar] [CrossRef]
- Schönswetter, P.; Schneeweiss, G.M. Is the incidence of survival in interior Pleistocene refugia (nunataks) underestimated? Phylogeography of the high mountain plant Androsace alpina (Primulaceae) in the European Alps revisited. Ecol. Evol. 2019, 9, 4078–4086. [Google Scholar] [CrossRef]
- Hampe, A.; Jump, A.S. Climate relicts: Past, present, future. Annu. Rev. Ecol. Evol. 2011, 42, 313–333. [Google Scholar] [CrossRef] [Green Version]
- Woolbright, S.A.; Whitham, T.G.; Gering, C.A.; Allan, G.J.; Bailey, J.K. Climate relicts and their associated communities as natural ecology and evolution laboratories. Trends Ecol. Evol. 2014, 29, 406–416. [Google Scholar] [CrossRef]
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.-G.; Cosson, J.-F. Comparative phylogeography and postglacial colonization routes in Europe. Mol. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Brochmann, C.; Gabrielsen, T.M.; Nordal, I.; Landvik, J.Y.; Elven, R. Glacial survival or tabula rasa? The history of North Atlantic biota revisited. Taxon 2003, 52, 417–450. [Google Scholar]
- Tribsch, A.; Schönswetter, P. Patterns of endemism and comparative phylogeography confirm palaeoenvironmental evidence for Pleistocene refugia in the eastern Alps. Taxon 2003, 52, 477–497. [Google Scholar] [CrossRef] [Green Version]
- Holderegger, R.; Thiel-Egenter, C. A discussion of different types of glacial refugia used in mountain biogeography and phylogeography. J. Biogeogr. 2009, 36, 476–480. [Google Scholar] [CrossRef]
- Berthouzoz, M.; Maendly, S.; Bétrisey, S.; Mangili, S.; Prunier, P.; Lexer, C.; Kozlowski, G. Some like it cold: Distribution, ecology and phylogeny of Arenaria bernensis Favarger (Caryophyllaceae) from the Western Prealps in Switzerland. Alp. Bot. 2013, 123, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Windmaisser, T.; Kattari, S.; Heubl, G.; Reisch, C. Glacial refugia and postglacial expansion of the alpine-prealpine plant species Polygala chamaebuxus. Ecol. Evol. 2016, 6, 7809–7819. [Google Scholar] [CrossRef] [PubMed]
- Stehlik, I.; Blattner, F.; Holderegger, R.; Bachmann, K. Nunatak survival of the high alpine plant Eritrichium nanum (L.) Gaudin in the Central Alps during the ice ages. Mol. Ecol. 2002, 11, 2027–2036. [Google Scholar] [CrossRef] [PubMed]
- Stehlik, I. Resistance or emigration? Response of alpine plants to the ice ages. Taxon 2003, 52, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Paun, O.; Schönswetter, P.; Winkler, M.; Consortium, I.; Tribsch, A. Historical divergence vs. contemporary gene flow: Evolutionary history of the calcicole Ranunculus alpestris group (Ranunculaceae) in the European Alps and the Carpathians. Mol. Ecol. 2008, 17, 4263–4275. [Google Scholar] [CrossRef]
- Ronikier, M.; Schneeweis, G.M.; Schönswetter, P. The extreme disjunction between Beringia and Europe in Ranunculus glacialis sl. (Ranunculaceae) does not coincide with the deepest genetic split—A story of the importance of temperate mountain ranges in arctic-alpine phylogeography. Mol. Ecol. 2012, 21, 5561–5578. [Google Scholar] [CrossRef]
- Winkler, M.; Tribsch, A.; Schneeweis, G.M.; Brodbeck, S.; Gugerli, F.; Holderegger, R.; Abbott, R.J.; Schönswetter, P. Tales of the unexpected: Phylogeography of the arctic-alpine model plant Saxifraga oppositifolia (Saxifragaceae) revisited. Mol. Ecol. 2012, 21, 4618–4630. [Google Scholar] [CrossRef]
- Eidesen, P.B.; Ehrich, D.; Bakkestuen, V.; Alsos, I.G.; Gilg, O.; Taberlet, P.; Brochmann, C. Genetic roadmap of the Arctic: Plant dispersal highways, traffic barriers and capitals of diversity. New Phytol. 2013, 200, 898–910. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, T.; Muster, C.; Schönswetter, P. Are Disjunct Alpine and Arctic-Alpine Animal and Plant Species in the Western Palearctic Really “Relics of the Cold Past”; Habel, J.C., Assman, T., Eds.; Relict Species: Phylogeny and Conservation Biology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 239–252. [Google Scholar]
- Zimmermann, M.; Vischer-Leopold, M.; Ellwanger, G.; Ssymak, A.; Schröder, E. The EU Habitat Directive and the German Natura 2000 Network of Protected Areas as Tool for Implementing the Conservation of Relict Species; Habel, J.C., Assman, T., Eds.; Relict Species: Phylogeny and Conservation Biology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 323–340. [Google Scholar]
- Parisod, C.; Besnard, G. Glacial in situ survival in the Western Alps and polytopic autopolyploidy in Biscutella laevigata L. (Brassicaceae). Mol. Ecol. 2007, 16, 2755–2767. [Google Scholar] [CrossRef] [Green Version]
- Hess, H.E.; Landolt, E.; Hirzel, R. Flora der Schweiz und Angrenzender Gebiete; Birkhäuser Verlag: Basel, Switzerland, 1977; Volume 2, p. 955. [Google Scholar]
- Richard, J.-L. La végétation du Vanil Noir et du Vallon des Morteys. Bul. Soc. Frib. Sc. Nat. 1977, 66, 1–52. [Google Scholar]
- Aeschimann, D.; Lauber, K.; Moser, D.M.; Theurillat, J.-P. Flora Alpina. Band 1–3; Haupt: Bern, Switzerland, 2004. [Google Scholar]
- Gerber, E.; Kozlowski, G.; Mariéthoz, A. La flore des Préalpes du lac de Thun au Léman; Rossolis: Bussigny, Switzerland, 2010. [Google Scholar]
- Kadereit, J.W.; Schwarzbach, A.E.; Jork, K.B. The phylogeny of Papaver s.l. (Papaveraceae): Polyphyly or monophyly? Plant Syst. Evol. 1997, 204, 75–98. [Google Scholar] [CrossRef]
- Carolan, J.C.; Hook, I.L.I.; Chase, M.W.; Kadereit, J.W.; Hodkinson, T.R. Phylogenetics of Papaver and related genera based on DNA sequences from ITS nuclear ribosomal DNA and plastid trnL intron and trnL-F intergenic spacers. Ann. Bot. 2006, 98, 141–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordal, I.; Hestmark, G.; Solstad, H. Reproductive biology and demography of Papaver radicatum—A key species in Nordic plant geography. Opera Bot. 1997, 132, 77–87. [Google Scholar]
- Solstad, H.; Elven, R.; Nordal, I. Isozyme variation among and within North Atlantic species of Papaver sect. Meconella (Papaveraceae) and taxonomic implications. Biol. J. Linn. Soc. 2003, 143, 255–269. [Google Scholar] [CrossRef]
- Kadereit, J.W. Notes on the taxonomy, distribution, phylogeny and ecology of Papaver alpinum L. (Papaveraceae). Bot. Jahrb. Syst. 1990, 112, 79–97. [Google Scholar]
- Schönswetter, P.; Solstad, H.; Escobar, G.P. A combined molecular and morphological approach to the taxonomically intricate European mountain plant Papaver alpinum s.l. (Papaveraceae)—Taxa or informal phylogeographical groups? Taxon 2009, 58, 1326–1348. [Google Scholar] [CrossRef]
- Markgraf, F. Papaveraceae. In Illustrierte Flora von Mitteleuropa, 2nd ed.; Hegi, G., Ed.; Hanser: München, Germany, 1958; Volume IV, pp. 15–49. [Google Scholar]
- Markgraf, F. Eine neue Gliederung der Alpenmohne. Phyton 1958, 7, 302–314. [Google Scholar]
- Pittet, L.; Fragnière, Y.; Grünig, S.; Bétrisey, S.; Clément, B.; Gerber, E.; Ronikier, R.; Kozlowski, G.; Parisod, C. Genetic structure of the narrow endemic Papaver occidentale indicates complex patterns of survival and immigration in the Western Prealps. Alp. Bot. 2020. [Google Scholar] [CrossRef]
- Bornand, C.; Gygax, A.; Juillerat, P.; Jutzi, M.; Möhl, A.; Rometsch, S.; Sager, L.; Santiago, H.; Eggenberg, S. Rote Liste Gefässpflanzen. Gefährdete Arten der Schweiz. Umwelt-Vollzug Nr. 1621; Bundesamt für Umwelt: Bern, Switzerland, 2016. [Google Scholar]
- Jordan, D. La Flore Rare ou Menacée de Haute-Savoie; Naturalia Publications: Turriers, France, 2015. [Google Scholar]
- Bornand, C.; Hoffer-Massard, F. La Pierreuse ou le parfum du pavot, le 20 juillet 2002. Bull. Cercle Vaud. Bot. 2003, 32, 53–61. [Google Scholar]
- Delarze, R. Plan d’action pour Papaver occidentale (Markgraf) H.E. Hess & Landolt (Pavot des Alpes occidentales); Coordination Régionale pour la Protection de la flore: Lausanne, Switzerland, 2012. [Google Scholar]
- Bétrisey, S. Adaptation du Plan d’action Pour Papaver Occidentale; Jardin botanique de l’Université de Fribourg: Fribourg, Switzerland, 2014. [Google Scholar]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions (MPB-49); Princeton University Press: Princeton, NJ, USA, 2011; Volume 56. [Google Scholar]
- Peterson, A.T.; Soberón, J. Species distribution modeling and ecological niche modeling: Getting the concepts right. Nat. Conserv. 2012, 10, 102–107. [Google Scholar] [CrossRef]
- Song, Y.-G.; Petipierre, B.; Deng, M.; Wu, J.-P.; Kozlowski, G. Predicting climate change impacts on the threatened Quercus arbutifolia in montane cloud forests in southern China and Vietnam: Conservation implications. For. Ecol. Manag. 2019, 444, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Buri, A.; Grand, S.; Yashiro, E.; Adatte, T.; Spangenberg, J.E.; Pinto-Figueroa, E.; Verrecchia, E.; Guisan, A. What are the most crucial soil variables for predicting the distribution of mountain plant species? A comprehensive study in the Swiss Alps. J. Biogeogr. 2020, 47, 1143–1153. [Google Scholar] [CrossRef]
- Hageer, Y.; Esperón-Rodríguez, M.; Baumgartner, J.B.; Beaumont, L.J. Climate, soil or both? Which variables are better predictors of the distributions of Australian shrub species? PeerJ 2017, 5, e3446. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Braunisch, V.; Coppes, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from correlated climate variables: A major source of uncertainty for predicting species distributions under climate change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Júnior, P.D.M.; Nóbrega, C.C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PLoS ONE 2018, 13, e0202403. [Google Scholar]
- Craney, T.A.; Surles, J.G. Model-dependent variance inflation factor cutoff values. Qual. Eng. 2002, 14, 391–403. [Google Scholar] [CrossRef]
- O’Brien, R.M. A caution regarding rules of thumb for variance inflation factors. Qual. Quant. 2007, 41, 673–690. [Google Scholar] [CrossRef]
- Neteler, M.; Mitasova, H. Open Source GIS: A GRASS GIS Approach; Springer Science & Business Media: Berlin, Germany, 2013; Volume 689. [Google Scholar]
- Menard, S. Applied Logistic Regression Analysis; Sage: Los Angeles, CA, USA, 2002; Volume 106. [Google Scholar]
- Wisz, M.S.; Guisan, A. Do pseudo-absence selection strategies influence species distribution models and their predictions? An information-theoretic approach based on simulated data. BMC Ecol. 2009, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- CH2018 Climate Scenarios for Switzerland; Technical Report; National Centre for Climate Services: Zurich, Switzerland, 2018; pp. 1–271.
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5. [Google Scholar] [CrossRef]
- Swisstopo. Metadata of the GeoCover V2 Product. 2018. Available online: https://shop.swisstopo.admin.ch/en/products/height_models/alti3D (accessed on 2 March 2020).
- MeteoSwiss. Monthly and Annual Reports. Federal Office of Meteorology and Climatology. Available online: https://www.meteoswiss.admin.ch (accessed on 2 March 2020).
- Swisstopo. Bundesamt für Landestopografie swisstopo-swissALTI3D-Das hoch aufgelöste Terrainmodell der Schweiz. Available online: https://shop.swisstopo.admin.ch/en/products/maps/geology/GC_VECTOR (accessed on 2 March 2020).
- McCullagh, P. Generalized Linear Models; Routledge: London, UK, 2019. [Google Scholar]
- Guisan, A.; Edwards, T.C., Jr.; Hastie, T. Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Hastie, T.J. Generalized additive models. In Statistical Models in S; Routledge: London, UK, 2017; pp. 249–307. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC: London, UK, 2017. [Google Scholar]
- Atkinson, A.C. A note on the generalized information criterion for choice of a model. Biometrika 1980, 67, 413–418. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Ishiguro, M.; Kitagawa, G. Akaike Information Criterion Statistics; D. Reidel 81: Dordrecht, The Netherlands, 1986. [Google Scholar]
- Rodriguez, J.D.; Perez, A.; Lozano, J.A. Sensitivity analysis of k-fold cross validation in prediction error estimation. IEEE Trans. Pattern Anal. Mach. Intell. 2009, 32, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Lever, J.; Krzywinski, M.; Altman, N. Points of significance: Model selection and overfitting. Nat. Methods 2016, 13, 703–704. [Google Scholar] [CrossRef]
- Shao, J. Linear model selection by cross-validation. J. Am. Stat. Assoc. 1993, 88, 486–494. [Google Scholar] [CrossRef]
- Xu, Q.S.; Liang, Y.Z. Monte Carlo cross validation. Chemomet. Intell. Lab. Syst. 2001, 56, 1–11. [Google Scholar] [CrossRef]
- Pohjankukka, J.; Pahikkala, T.; Nevalainen, P.; Heikkonen, J. Estimating the prediction performance of spatial models via spatial k-fold cross validation. Int. J. Geogr. Inf. Sci. 2017, 31, 2001–2019. [Google Scholar] [CrossRef]
- Roberts, D.R.; Bahn, V.; Ciuti, S.; Boyce, M.S.; Elith, J.; Guillera-Arroita, G.; Hauenstein, S.; Lahoz-Monfort, J.J.; Schröder, B.; Thuiller, W.; et al. Cross-validation strategies for data with temporal, spatial, hierarchical, or phylogenetic structure. Ecography 2017, 40, 913–929. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org (accessed on 2 March 2020).
- QGIS Development Team. QGIS Geographic Information System; Open Source Geospatial Foundation Project: Chicago, IL, USA, 2019; Available online: https://www.qgis.org/fr/site (accessed on 2 March 2020).
- Petrovsky, V.V. The problem of the species-width in poppies of Russian Arctic. Skr. Norske Vidensk. Akad. Oslo 1999, 38, 243–259. [Google Scholar]
- PAF. Panarctic Flora Project—Annotated Checklist of the Panarctic Flora–Vascular Plants. 2018. Available online: http://panarcticflora.org (accessed on 30 January 2020).
- Solstad, H.; Elven, R.; Nordal, I. Are there too many species and subspecies in the Papaver radicatum complex? Skr. Norske Vidensk. Akad. Oslo 1999, 38, 281–294. [Google Scholar]
- Paus, A. Late Weichselian vegetation, climate, and floral migration at Sandvikvatn, North Rogaland, south-western Norway. Boreas 1988, 17, 113–139. [Google Scholar] [CrossRef]
- Dubiel, E.; Olech, M. Plant communities of NW Sörkapp Land (Spitsbergen). Zesz. Nauk. Uniw. Jagiellońskiego 1990, 21, 35–74. [Google Scholar]
- Dubiel, E. Ecological observations on vascular plants in the NW Sörkapp Land (Spitsbergen). Zesz. Nauk. Uniw. Jagiellońskiego 1991, 22, 39–46. [Google Scholar]
- Dubiel, E. Geobotanical problems of NW Sörkapp Land (Spitsbergen). Zesz. Nauk. Uniw. Jagiellońskiego 1991, 22, 15–38. [Google Scholar]
- Thuiller, W.; Lavorel, S.; Araujo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Guerrina, M.; Conti, E.; Minuto, L.; Casazza, G. Knowing the past to forecast the future: A case study on a relictual, endemic species of the SW Alps. Berardia Subacaulis Reg. Environ. Chang. 2016, 16, 1035–1045. [Google Scholar] [CrossRef]
- Casazza, G.; Giordani, P.; Benesperi, R.; Foggi, B.; Viciani, D.; Filigheddu, R.; Farris, E.; Bagella, S.; Pisanu, S.; Mariotti, M.G. Climate change hastens the conservation urgency of range-restricted plant species in the central-northern Mediterranean region. Biol. Conserv. 2014, 179, 129–138. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum eleveation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Dulliger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plant under twenty-first-century climate change. Nat. Clim. Chang. 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Cotto, O.; Wessely, J.; Georges, D.; Klonner, G.; Schmid, M.; Dulliger, S.; Thuiller, W.; Guillaume, F. A dynamic eco-evolutionary model predicts slow response of alpine plants to climate warming. Nat. Commun. 2017, 8, 15399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, J.M.; Chalmandrier, L.; Lenoir, J.; Burgess, T.I.; Essl, F.; Haider, S.; Kueffer, C.; McDougall, K.; Milbau, A.; Nunez, M.A.; et al. Lags in the response of mountain plant communities to climate change. Glob. Chang. Biol. 2017, 24, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Patsiou, T.S.; Conti, E.; Zimmermann, N.E.; Theodoridis, S.; Randin, C. Topo-climatic microrefugia explain the persistence of a rare endemic plant in the Alps during the last 21 millennia. Glob. Chang. Biol. 2014, 20, 2286–2300. [Google Scholar] [CrossRef] [PubMed]
- Suchan, T.; Malicki, M.; Ronikier, M. Relict populations and Central European glacial refugia: The case of Rhododendron ferrugineum (Ericaceae). J. Biogeogr. 2019, 46, 392–404. [Google Scholar] [CrossRef]
- Vittoz, P.; Randin, C.; Dutoit, A.; Bonnet, F.; Hegg, O. Low impact of climate change on subalpine grassland in the Swiss Northern Alps. Glob. Chang. Biol. 2009, 15, 209–220. [Google Scholar] [CrossRef]
- Aubert, S. La flore du Mont Tendre en raccourci. Bull. Cercle Vaud. Bot. 1952, 2, 25–26. [Google Scholar]
- Moret, J.-L. Les plantes introduites à la Valée de Joux. Bull. Cercle Vaud. Bot. 2008, 37, 95–105. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Variable Type | Unit | Source | Native Resolution | Resolution Used in SDM |
|---|---|---|---|---|---|
| ELEVATION | Digital elevation model (DEM) | Meters (m) | swissALTI3D, The high precision digital elevation model of Switzerland [59] | 2 m, vertical uncertainty ±0.5 m < 2000 m, 1–3 m > 2000 m | Upscaled to 10 m, median value |
| SLOPE | terrain inclination | Degrees (°) | Calculated from DEM | 2 m | Upscaled to 10 m, median value |
| NORTHNESS | Slope aspect on the south-north axis | No unit, a real number between −1 (south) and 1 (north) | Calculated from DEM, cosinus of the slope aspect | 2 m | Upscaled to 10 m, median value |
| EASTNESS | Slope aspect on the west-east axis | No unit, a real number between −1 (west) and 1 (east) | Calculated from DEM, sinus of the slope aspect | 2 m | Upscaled to 10 m, median value |
| TEMP | Mean yearly mean temperature (norm, 1981–2010) | degrees °C | TnormY8110 [59] | 1000 m | Downscaled to 10 m, using DEM to improve resolution (strong negative linear relationship between ELEVATION and TEMP, R-squared value = 0.92, p-value < 0.001. TEMP was thus calculated as a function of ELEVATION, see Table S2, Figure S1, Supplementary Materials) |
| TEMP_Sum | Mean summer mean temperature (norm, 1981–2010) | degrees °C | TnormM8110 mean for June, July and August [59] | 1000 m | Downscaled to 10 m, same procedure as TEMP |
| TEMP_Win | Mean winter mean temperature (norm, 1981–2010) | degrees °C | TnormM8110, mean for December, January and February [59] | 1000 m | Downscaled to 10 m, same procedure as TEMP |
| PREC | Mean yearly precipitation (norm, 1981–2010) | Millimeters (mm) | RnormY8110 [59] | 1000 m | Downscaled to 10 m, using bicubic spline interpolation to get a smooth result (avoid artificial steps) |
| PREC_Sum | Mean summer precipitation (norm, 1981–2010) | Millimeters (mm) | RnormM8110, mean for June, July and August [59] | 1000 m | Downscaled to 10 m, using bicubic spline interpolation to get a smooth result (avoid artificial steps) |
| PREC_Win | Mean winter precipitation (norm, 1981–2010) | Millimeters (mm) | RnormM8110, mean for December, January and February [59] | 1000 m | Downscaled to 10 m, using bicubic spline interpolation to get a smooth result (avoid artificial steps) |
| SUNSHINE | Mean yearly relative sunshine duration (norm, 1981–2010) | Percent (%) | SnormY8110 [59] | 1000 m | Downscaled to 10 m, using bicubic spline interpolation to get a smooth result (avoid artificial steps) |
| IRRAD | Total irradiance during one day at the solstice (21st June) | Wh/m2/day | Calculate from DEM with r.sun (Solar irradiance and irradiation model) in GRASS GIS [60] | 10 m | 10 m |
| HOURS_SUN | Hours of sun during one day at the solstice (21st June) | Hours (h) | Calculate from DEM with r.sun (Solar irradiance and irradiation model) in GRASS GIS [60] | 10 m | 10 m |
| SCREES | Landcover: area with presence of calcareous screes | Presence-absence (1–0) | Area delimited using GEOCOVER (Geological vector datasets for better subsurface management, [61]) and interpretation of orthophotos (www.swisstopo.ch, Google maps). A 100 m buffer was added to the delimited areas, as the precise limit of the scree is difficult to get. | NA, vector data. | Rasterized to 10 m |
| Climatic Change Scenario | Remaining Suitable Area (Entire Studied Area) |
|---|---|
| Actual | 100% |
| RCP 2.6, year 2085 | 30% |
| RCP 4.5, year 2085 | 4% |
| RCP 8.5, year 2085 | 0% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fragnière, Y.; Pittet, L.; Clément, B.; Bétrisey, S.; Gerber, E.; Ronikier, M.; Parisod, C.; Kozlowski, G. Climate Change and Alpine Screes: No Future for Glacial Relict Papaver occidentale (Papaveraceae) in Western Prealps. Diversity 2020, 12, 346. https://doi.org/10.3390/d12090346
Fragnière Y, Pittet L, Clément B, Bétrisey S, Gerber E, Ronikier M, Parisod C, Kozlowski G. Climate Change and Alpine Screes: No Future for Glacial Relict Papaver occidentale (Papaveraceae) in Western Prealps. Diversity. 2020; 12(9):346. https://doi.org/10.3390/d12090346
Chicago/Turabian StyleFragnière, Yann, Loïc Pittet, Benoît Clément, Sébastien Bétrisey, Emanuel Gerber, Michał Ronikier, Christian Parisod, and Gregor Kozlowski. 2020. "Climate Change and Alpine Screes: No Future for Glacial Relict Papaver occidentale (Papaveraceae) in Western Prealps" Diversity 12, no. 9: 346. https://doi.org/10.3390/d12090346