

Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania

Nature Research Centre, Institute of Botany, Žaliųjų Ežerų Str. 49, 12200 Vilnius, Lithuania
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(1), 107; https://doi.org/10.3390/d15010107
Submission received: 5 November 2022
/
Revised: 9 January 2023
/
Accepted: 10 January 2023
/
Published: 12 January 2023
(This article belongs to the Special Issue Changes and Evolution of Flora and Vegetation under Human Impacts)
Abstract
:Most alien plant species are usually discovered in new areas either immediately or within a relatively short period of time after their introduction. However, the introduction, spread, and invasion of some species go unnoticed for a long time. In Lithuania, such an alien plant is Cornus sanguinea subsp. australis, whose cryptic invasion has gone unnoticed due to its similarity to the native subsp. sanguinea. Cornus sanguinea subsp. australis is native to southeastern Europe and western Asia and is considered alien in other parts of Europe. This study, which was carried out between 2018 and 2022 in the territory of Lithuania, aimed to identify the distribution of the subsp. australis, as well as assess certain reproductive traits and its impact on the species diversity of the invaded communities. The results showed that the cryptic invasion of subsp. australis in Lithuania has been going on for several decades and is now well advanced. We found that the alien subspecies grows faster and that individuals in analogous habitats mature earlier than the native subsp. sanguinea. Furthermore, subsp. australis, compared to subsp. sanguinea, produces more flowers per inflorescence (84.0 ± 25.0 and 49.7 ± 20.9, respectively) and sets and ripens more fruits (25.6 ± 9.7 and 18.2 ± 7.6, respectively), which are among the most important factors for its further fast spread and invasion. The results of the vegetation analysis showed that the increase in abundance of both subspecies has a negative effect on species diversity in the community. However, the effect of the native subspecies on plant species diversity was negligible, while the effect of the alien subspecies was moderately negative and significant (r = −0.47; p < 0.05). Based on the results of the study, we can conclude that the alien subsp. australis performs better than the native subsp. sanguinea. Considering the current distribution and the drivers of dispersal, we suppose that subsp. australis will continue to spread rapidly in Lithuania and may pose a threat to the native subsp. sanguinea populations.
1. Introduction
Most invasions of alien plant species into new areas are detected as soon as they begin, and the whole process is usually well documented [1,2,3]. However, there are some alien plants whose introduction, naturalization, and spread in new areas go unnoticed for decades because of their similarity to native or widespread alien plants [4]. The invasion of alien species, taxa of other ranks, races, or genotypes that are morphologically close to well-known organisms, and therefore go undetected for long periods of time, is known as cryptic invasion [5,6,7,8].
Cryptic plant invasions are often the result of the introduction of alien infraspecific taxa that closely resemble native taxa, as well as alien races or genotypes that can only be reliably identified using molecular techniques [7,9]. The spread of some infraspecific taxa (or races) of Phragmites australis (Cav.) Trin. ex Steud. in different regions of the world can be considered a cryptic invasion of the largest scale. Some alien infraspecific taxa of this species have been found to be significantly different from native ones in relation to their rates of spread, occupied habitats, and impact on ecosystems [8]. Studies on the cryptic invasion of Ceratophyllum demersum L. in New Zealand have revealed that its alien populations are highly invasive and behave differently from native populations [10]. Recently, the alien Hedera hibernica Poit. has been found to be spreading particularly fast in Belgium, but it is difficult to distinguish it from the native Hedera helix L. by its morphological characteristics. Hedera hibernica grows much more vigorously than Hedera helix and occupies habitats unsuitable for the native species [11]. Thus, because of the cryptic nature of certain invasive species, they can spread uncontrolled for long periods of time, causing a detrimental impact on native species and ecosystems. Once an invasion has become apparent, it may already be too late to control the spread of the invasive species, manage it effectively, or mitigate the adverse effects of the invasion [7].
The invasiveness of a plant species is determined by many environmental factors and plant traits [12,13]. The most important factors determining the invasiveness of woody plants include their fleshy fruits (which contain seeds distributed by birds), short juvenile periods (less than 10 years), and short intervals (1 to 4 years) between large seed crops [14]. The duration of the juvenile period of woody plants can be determined not only under experimental conditions but also from annual rings. Dendrochronological methods can also be used to determine the age structure of woody plant populations with a reasonable degree of precision, as well as the time of the establishment of individuals in an area, the age of maturity, and the rate of spread or growth [15,16,17]. Despite the simplicity of the dendrochronological method, it is still used relatively rarely in invasive species studies [18,19].
Forestry, landscape, and ornamental planting are major contributors to the increase, spread, and invasion of alien woody plant species worldwide [20,21]. Studies have shown that, the greater the intensity of the introduction, the heavier the propagule pressure, which facilitates the spread of the alien species and increases the likelihood of its invasion [22]. Elaeagnus angustifolia L. has been planted as an ornamental and sheltering species for a long time over a large area in Canada and the United States but is now an invasive plant with a detrimental impact on riparian ecosystems [23]. Recently, there has been an increasing emphasis on the significance of shelterbelts and hedgerows composed of alien woody plants along streets, roads, railways, or other infrastructure as a reservoir for invasive and potentially invasive species [24,25,26].
One of the most important objectives of research on invasive species is to determine the consequences of invasion at different scales from the impacts on individual species and changes in ecosystem functioning [27,28]. Most knowledge has been gained on the competition of invasive species with native species and the resulting changes in plant communities [29,30]. Biodiversity is most affected when invasive species dominate communities, and such communities cover large areas [31]. Native species with strong competitive characteristics and dominance in communities are also known to reduce species diversity, but in such cases, the loss of diversity is less pronounced and is usually reversible [30,32].
The introduction of plants of the genus Cornus L. for ornamental purposes began around the 17th century in Europe and is still ongoing [33,34]. The spread from cultivation sites and the naturalization of some introduced species started soon after their introduction. The number of recorded alien species of the genus Cornus in various European countries is steadily increasing [34,35,36,37]. The most widespread and invasive species in Europe are now Cornus alba L. and Cornus sericea L. [38,39,40]. During several decades, parts of Europe have witnessed an intensive spread of the alien Cornus sanguinea subsp. australis (C.A. Mey.) Jáv. ex Soó into anthropogenic, semi-natural, and even natural habitats [41]. This subspecies has been and continues to be planted in protective and ornamental plantations [42].
The alien subsp. australis has been growing in Lithuania for at least eight decades but has only recently been identified. The plant has become fully naturalized during this time and has become widespread in the country [37]. We, therefore, undertook a study to assess the extent of the cryptic invasion of subsp. australis, and to compare its biological and ecological characteristics with the native subsp. sanguinea. In this study, we aimed to answer the following questions: (i) How widely is subsp. australis distributed in Lithuania and what are the factors underlying its cryptic invasion? (ii) Are there differences between the reproductive characteristics of alien subsp. australis and native subsp. sanguinea? (iii) Are there differences in the age of maturity of young individuals of the two subspecies? (iv) What are the effects of both subspecies on the species diversity of plant communities?
2. Materials and Methods
2.1. Study Taxa
Cornus sanguinea subsp. australis (C.A. Mey.) Jáv. ex Soó is distinguished from the subsp. sanguinea by the type of hairs on the abaxial surface of its leaves. Cornus sanguinea subsp. australis has medifixed hairs appressed to the leaf surface, whereas the subsp. sanguinea has simple, curved loose hairs [43]. Discussions on the rank of this taxon have continued for decades [44,45], and even now some authors treat this taxon differently. Woźnicka et al. (2015) [46] and Morozowska et al. (2021) [47] showed that there are no significant differences in the morphological features of the external and internal structure of the endocarp of subsp. sanguinea and subsp. australis. Nevertheless, sometimes this subspecies is treated at the rank of species, i.e., Cornus australis C.A. Mey. [43]. Following recent studies [46,47,48], we accept these two taxa at the rank of subspecies.
Cornus sanguinea subsp. australis is a shrub or small tree native to southeastern Europe and western Asia [48,49,50]. In central and western Europe (e.g., Belgium, the Netherlands, Germany, and Austria), it is treated as an alien [41,42,43,51,52]. In the northeastern part of Ukraine, as well as Lithuania and Latvia, subsp. australis is also considered alien [37,53,54]. In its native range, subsp. australis is mostly associated with the woodlands of the forest-steppe zone, sparse woodlands, forest edges, and slopes of mountain valleys [49,50].
2.2. Distribution
The distribution of subsp. australis in Lithuania was studied from 2018 to 2022. The search for plants of this taxon was carried out primarily in areas where they were most likely to be found, i.e., in urbanized areas and along major railways, as well as in natural and semi-natural habitats at varying distances from the established populations of the subsp. australis. A revision of all specimens of Cornus sanguinea deposited in the herbaria of Nature Research Centre (BILAS) and Vilnius University (WI) was performed to establish the historical records of this subspecies. Herbarium specimens collected during this research were deposited at the Herbarium of the Nature Research Centre in Vilnius. A total of 48 herbarium specimens were examined and more than 50 specimens were collected during the research. Plants were identified by morphological characteristics, the most important of which was the type of hairs on the underside of the leaves [43,48].
The introduction pathways and vectors of spread were defined and classified according to guidelines for the prioritization and management of invasive species [55].
A distribution map of subsp. australis in Lithuania was compiled, applying a system of grid cells that were arranged according to the geographical coordinates with sides of 6’ of latitude and 10’ of longitude [56]. Sites recorded in the same grid cell were marked on the map with a single dot.
2.3. Study of Plant Communities
Phytosociological surveys of the plant communities inhabited by both subspecies of Cornus sanguinea were carried out throughout Lithuania. In selecting the study areas, we tried to cover as much habitat diversity as possible, considering the abundance of the taxa studied. In assessing the presence of communities with a different abundance of subsp. australis and subsp. sanguinea, we wanted to assess their effect on species richness. Community composition and species coverage were assessed in 100 m2 study plots selected in a homogeneous part of the habitat [57]. The coverage of each plant species was visually assessed and expressed as a percentage. The coverage of each plant species in different layers (trees, shrubs, herbs, and mosses) was assessed separately, in addition to the total coverage of each layer.
In total, 60 plots were used for the analysis (30 communities with subsp. australis and 30 communities with subsp. sanguinea). All communities with plants of each Cornus sanguinea subspecies were divided into two groups. Communities with more than 40% tree cover were considered woodlands, and communities with less than 40% tree cover were considered shrublands.
2.4. Study of Reproductive Traits
Aiming to assess the reproductive traits of subsp. australis and subsp. sanguinea, studies were performed in three populations of each subspecies in 2020 (Figure 1, Table 1). For this study, populations of both subspecies were selected from similar habitats. The Cornus sanguinea subsp. sanguinea reproductive characteristics were studied in the habitats of shrublands (Piepaliai and Veliuona sites) and Quercus robur woodland (Šilinė site), while subsp. australis populations were selected in anthropogenic woodland (Kaišiadorys site) and abandoned grassland (Maišiagala and Ginkūnai sites) habitats.
In each population, three inflorescences (one inflorescence each from the lower, middle, and upper branches) were collected from ten tagged individuals during the flowering period (early June). The number of all normally developed flowers in each inflorescence was counted. In total, 30 inflorescences were analyzed in each population. At the end of the summer, three infructescences were collected from the same ten labeled individuals in each population. The number of all normally developed fruits per infructescence was counted. In total, 30 infructescences were examined in each population. The sample size for estimating the number of flowers in an inflorescence was determined according to the results of the pilot study and following the recommendations by Lenth [58]. The same sample size was applied to assess the number of fruits per infructescence.
2.5. Age of Maturity
To determine whether there were differences in the age of maturity of subsp. australis and subsp. sanguinea, we carried out dendrochronological studies on young but already fruiting shoots, presumably grown from root suckers. At four sites (two subsp. australis sites and two subsp. sanguinea sites), 30 of the youngest fruiting shoots were collected in September 2020. The fruit-bearing shoots were selected if they were 1.0–1.1 m tall or less. In Vievis (subsp. australis; 54.77093° N; 24.83455° E) and Piepaliai (subsp. sanguinea; 55.07433° N; 23.78359° E), the samples were collected in xeromesic habitats, and in Kaišiadorys (subsp. australis; 54.87055° N; 24.43644° E) and Veliuona (subsp. sanguinea; 55.08996° N; 23.18326° E), the samples were collected in mesic habitats. All habitats were open, without shade from other trees and shrubs. All, or at least most, of the shoots sampled were presumably derived from root suckers.
The shoots were cut with secateurs at the soil surface, a section of the lower part of about 15 cm was taken, and each sample was numbered. In the laboratory, a section of the shoots was examined, and the age of the shoot was determined by the number of annual rings. The annual rings were counted by examining the section under a binocular microscope. The diameter of each shoot was measured with a caliper (accuracy 0.1 mm) at its base.
2.6. Statistical Analyses
The evaluation of the collected research data using the Shapiro–Wilk test showed that some of the data on the number of flowers per inflorescence, the number of fruits per infructescence, the age of the young individuals and their stem diameter, as well as the number of species in the studied plant communities, did not meet the criteria for normal distribution. Therefore, we compared the datasets using non-parametric statistical analysis methods. The Kruskal–Wallis H-test to detect differences between datasets from all study sites, and the Mann–Whitney U-test for post hoc pairwise comparison was applied. To compare the number of flowers and fruits of the taxa under study, the data from all study sites were pooled and the resulting datasets were compared with each other. When presenting the results of the descriptive statistics, we report the mean and the standard deviation (mean ± SD), and when considering the age and stem diameter of young individuals, we also report the minimum, maximum (min–max), and median values. A nested two-way ANOVA was used to assess the effect of the taxon (fixed effect) and habitat (random effect) on age at the start of shoot fruiting. The relationships between the cover of the studied Cornus taxa and plant species diversity in the communities were assessed using Spearman’s rank correlation (rs). For the comparison of significant differences between the samples from the literature and our data, we used the t-test from parameters. All the calculations were performed using the PAST 4.10 software [59].
3. Results
3.1. Distribution
Examination of the herbarium specimens of Cornus sanguinea from Lithuania revealed that the oldest sample of the subsp. australis was collected by P. Snarskis on 26 July 1940 (BILAS, No. 6549, originally determined as Cornus sanguinea) on the western side of the narrow-gauge railway line Panevėžys–Joniškėlis, ca. 4 km north of Bernatoniai (now in the Panevėžys district) railway station (Figure 2). However, P. Snarskis did not indicate on the herbarium label whether the specimen represented a cultivated or a self-grown individual.
The results of the study showed that the subsp. australis was recorded in 63 grid cells in Lithuania, with a total area of occurrence of ca. 40,400 km2, comprising about 63% of the country’s territory (Figure 2). The total number of recorded localities of the subsp. australis in Lithuania in 2022 was estimated to be more than 200, as one to ten sites were recorded in individual grid cells.
In the southern, central, and eastern parts of Lithuania, subsp. australis is relatively concentrated. In the northern and western parts of the country, the occurrence of the subspecies is more diffuse, and in the southwestern part of the country, the subspecies has not yet been recorded. In most cases, subsp. australis has spread from road and rail shelter plantations, and less frequently from ornamental plantations. From shelter and ornamental plantations, these plants spread under the influence of anthropogenic and natural vectors. Among the anthropogenic vectors, the disposal of fallen leaves and other plant waste should be considered the most important, while the most important natural vector is fruit-feeding birds. The importance of birds for the spread of subsp. australis is demonstrated by the fact that, in many areas, the subspecies forms thickets of varying size underneath power lines and around electricity poles, on which flocks of frugivorous birds often land to take a rest.
Cornus sanguinea subsp. australis was observed in a variety of anthropogenic habitats (anthropogenic woodlands, roadsides, railways, wastelands, margins of drainage ditches, abandoned lands, urbanized areas, and their surroundings), but also often occurred in semi-natural or natural habitats. In natural and semi-natural habitats, subsp. australis was most frequently found along forest edges and in hillside grasslands, and less frequently on the banks of water bodies. Assessing the current distribution, extensive stands, and dispersal vectors, it can be assumed that subsp. australis will continue to spread and invade anthropogenic, semi-natural, and natural habitats.
3.2. Comparison of Reproductive Traits of Subspecies
3.2.1. Flowering and Fruiting
Analysis of the number of flowers per inflorescence showed that the mean number of flowers per inflorescence of subsp. sanguinea (49.7 ± 20.9 flowers) was significantly lower (U = 1134; p < 0.001) than the mean number of flowers per inflorescence of subsp. australis (84.0 ± 25.0 flowers). The mean number of flowers per inflorescence at individual subsp. sanguinea sites ranged from 36.1 ± 14.5 to 65.7 ± 21.0, while the mean number at subsp. australis sites ranged from 78.2 ± 23.2 to 87.2 ± 23.5 flowers per inflorescence (Figure 3). The results of the Kruskal–Wallis test showed that among all studied sites of subsp. sanguinea and subsp. australis, significant differences (H = 91.88; p < 0.001) in the number of flowers per inflorescence were found. It should be noted that in terms of the number of flowers per inflorescence, we found no significant differences between subsp. sanguinea at the Piepaliai site and subsp. australis at the Ginkūnai site (U = 320; p = 0.05). A comparison of all three subsp. australis sites according to the number of flowers per inflorescence revealed no significant differences (H = 2.2; p = 0.33), while between all sites of subsp. sanguinea, significant differences (H = 30.82; p < 0.001) were found. This suggests that subsp. australis has a low response to environmental conditions, while the native subsp. sanguinea is only comparable to the alien subspecies in terms of the number of flowers per inflorescence when it grows under particularly favorable conditions, i.e., in open xerothermic habitats.
The results showed that, in subsp. sanguinea, infructescence mean fruit number at the study sites (18.2 ± 7.6 fruits) was significantly lower (U = 2095; p < 0.01) than the mean fruit number of subsp. australis (25.6 ± 9.7 fruits). The mean number of fruits per infructescence at individual subsp. sanguinea sites ranged from 36.1 ± 12.8 ± 22.2 to 21.9 ± 8.3, while the mean number at subsp. australis sites ranged from 23.3 ± 8.9 to 28.1 ± 10.8 fruits per infructescence (Figure 3). The Kruskal–Wallis test revealed that among all sites of subsp. sanguinea and subsp. australis, there were significant differences in the number of fruits per infructescence (H = 57.48; p < 0.001). Pairwise comparisons of sites by the number of fruits per infructescence showed slightly different patterns than comparisons by the number of flowers per inflorescence (Figure 3). At the Šilinė site, subsp. sanguinea produced significantly fewer fruits than all the other sites studied. Between the two subsp. sanguinea and two subsp. australis sites, the number of fruits in the infructescence did not differ significantly (Figure 3). A comparison of all three subsp. australis sites according to the number of fruits per infructescence revealed no significant differences (H = 4.4; p = 0.11), while significant differences were found between all sites of subsp. sanguinea (H = 29.41; p < 0.001).
The analysis of the research data showed that subsp. sanguinea (36.6% of the potential fecundity) had a higher realized fecundity than that of subsp. australis (30.5% of the potential fecundity). Nevertheless, inflorescences of subsp. australis had significantly more flowers than inflorescences of subsp. sanguinea (7558 and 4475 flowers in the 90 inflorescences of each subspecies analyzed, respectively), and significantly more fruit ripened in the infructescences (2304 and 1638 fruits, respectively). At individual sites, the realized fecundity of subsp. sanguinea ranged from 27.0% to 55.0% of the potential fecundity, while that value of subsp. australis ranged between 26.7% and 35.9%.
3.2.2. Maturation Age
Significant differences were found between sites in terms of the age of shoots starting to fruit by applying the Kruskal–Wallis H-test (H = 67.64, p < 0.001). A pairwise comparison showed that there were no significant differences in shoot age between sites of subsp. australis and sites of subsp. sanguinea (Table 2). However, both sites of subsp. australis were significantly different from both sites of subsp. sanguinea sites regarding the age of the shoots. It was found that subsp. australis shoots start fruiting at a significantly younger age than subsp. sanguinea (U = 254.0, p < 0.001). Analysis of the data by the diameter of the shoots at the beginning of fruiting (Table 2) showed significant differences between sites (H = 88.20, p < 0.001). Pairwise comparisons also showed significant differences between all sites. It was found that subsp. australis shoots start fruiting at a significantly smaller diameter than subsp. sanguinea (U = 79.0, p < 0.001).
The results of the two-way nested ANOVA analysis showed that taxon had a significant effect on the age of the root suckers that had started fruiting, while mesic and xeromesic habitat types did not have a significant effect on the age of fruiting. Shoot diameter was significantly influenced by both factors, taxon, and habitat. In mesic habitats, both species had a larger shoot diameter than in xeromesic habitats, irrespective of age (Table 3).
3.3. Species Diversity in Invaded Communities
A total of 30 study plots were analyzed for each subspecies: subsp. australis and subsp. sanguinea. These were divided into two groups: woodlands and shrublands. There were no significant differences between the cover of the taxa studied and the cover of trees, herbs, and bryophytes between the plots of the two subspecies. Total shrub cover was significantly higher in the study plots with native subsp. sanguinea (Table 4).
An examination of the composition of the communities with two subspecies of Cornus sanguinea found that in the study plots with alien subsp. australis, a total of 160 plant species were recorded, while in the plots with native subsp. sanguinea, 170 plant species were recorded. In the study plots with subsp. australis, the number of plant species ranged from 8 to 45, while the number of species in the study plots with subsp. sanguinea ranged from 11 to 36. The mean number of species in the study plot with subsp. australis was 22.6 ± 8.2 (median 21) species, while the number of species in plots with subsp. sanguinea was 24.6 ± 6.4 (median 24) species (Table 4). There were no significant differences in the number of species between the communities with the subspecies studied (U = 359.0; p = 0.129).
In the communities studied with subsp. australis, a total of 55 woody plant species were recorded, while in the communities with subsp. sanguinea, 33 such species were recorded. In the study plot, in the communities with subsp. australis, the mean number of species was 8.2 ± 4.3 (median 7), while the communities with subsp. sanguinea contained a mean of 7.0 ± 2.7 (median 7) species. There were no significant differences in the number of woody plant species between communities with the subspecies studied (U = 397.5; p = 0.331). Somewhat different patterns were found when examining the communities by herbaceous species composition. In the communities with alien subsp. australis, a total of 94 herbaceous plant species were recorded, while with the native subsp. sanguinea, there were 135 herbaceous plant species. In communities with subsp. australis, the mean number of species recorded in the study plot was 12.9 ± 6.8 (median 12), while in the study plot with subsp. sanguinea, the mean number of species was 15.8 ± 5.8 (median 16). It was found that in communities with subsp. australis, the diversity of herbaceous plant species was significantly lower (U = 323.0; p = 0.041) than in communities with subsp. sanguinea (Figure 4).
Communities with alien subsp. australis were more diverse than communities with native subsp. sanguinea when considering the number of alien species. In the study plots with subsp. australis, a total of 27 alien species were recorded, while in the study plots with subsp. sanguinea, 9 alien species were recorded. The mean number of alien species in the study plot was 3.5 ± 1.6 (median 3), while the mean number of alien species in the study plot with the native subspecies was 0.6 ± 1.1 (median 0). It was found that, in communities with subsp. australis, the diversity of alien plant species was significantly higher (U = 60.5; p < 0.001) than in communities with subsp. sanguinea (Figure 4). However, the overall frequency of alien species was low, with most of them being recorded in only one study plot. The most common alien species in communities with subsp. australis were Physocarpus opulifolius (L.) Maxim. (recorded in 26.7% of study plots), Acer negundo L. (16.7%), Amelanchier spicata (13.3%), Ligustrum vulgare (13.3%), and Impatiens parviflora DC. (13.3%). Nevertheless, in the individual study plots, alien species were quite abundant: Physocarpus opulifolius covered from 0.5% to 30% of the plot surface, Acer negundo cover ranged from 5% to 15%, and Impatiens parviflora covered from 10% to 25%. In communities with subsp. sanguinea, the most frequent alien species were Impatiens parviflora (recorded in 16.7% of study plots) and Ligustrum vulgare (10.0%). Impatiens parviflora cover in these communities ranged from 1% to 10%, and the cover of Ligustrum vulgare ranged from 0.5% to 1%.
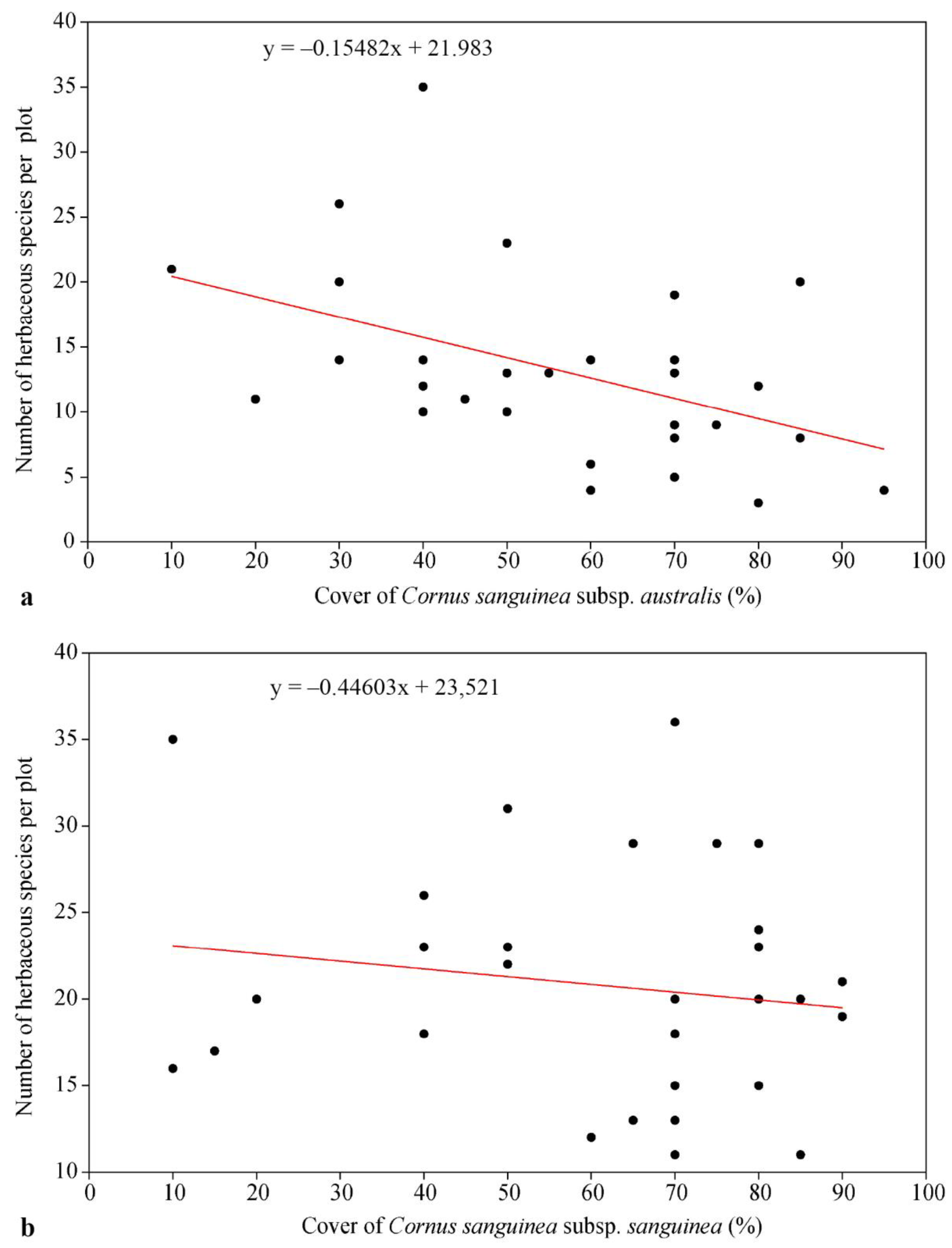
The correlation analysis showed that with an increase in the cover of alien subsp. australis, the diversity of herbaceous plant species decreased significantly (rs = −0.48; p = 0.007; r2 = 0.22; Figure 5a). In contrast, no significant decline in herbaceous plant species diversity was revealed in plant communities with an increase in cover of the native subsp. sanguinea (rs = −0.10; p = 0.575; r2 = 0.02; Figure 5b). It should be noted that there was no difference in cover between the two subspecies of Cornus sanguinea in the study plots (Table 4), but the mechanism of the effect of subsp. australis on herbaceous plant diversity is currently unclear.
The analysis of the most frequent plant species in communities with alien subsp. australis and native subsp. sanguinea revealed that both subspecies occupy similar woodland communities (Table 5). In shrubland communities with alien subsp. australis, species indifferent to environmental conditions had a higher frequency (e.g., Veronica chamaedrys L., Carex hirta L, Taraxacum officinale F.H. Wigg., and Equisetum arvense L.), while communities with native subsp. sanguinea shrubland communities had more species typical of mesic and xeromesic habitats among the most frequent species (e.g., Poa pratensis L., Agrimonia eupatoria L., Briza media L., and Medicago falcata L).
The results of the study suggest that the alien subsp. australis occupies habitats analogous or similar to those of the native subsp. sanguinea. The occupation of analogous habitats by both subspecies creates conditions for them to form mixed populations, leading to hybridization.
4. Discussion
4.1. Distribution
In Lithuania, the first herbarium specimen of Cornus sanguinea subsp. australis was collected more than eight decades ago, but it was not identified to the level of subspecies [37]. The spread of this subspecies has only recently been noticed, but the current distribution is the result of a cryptic invasion that has been ongoing for several decades. Based on the scarce data, we can assume that subsp. australis has been planted in shelter plantations along railways and roads in Lithuania since the 1940s, and possibly earlier. We have not been able to find any data on the origin of the seedlings used for planting or the seeds from which the seedlings were grown. It is reported that, for a long time, subsp. australis was also planted in other European countries along railways and roads (e.g., in Belgium, Germany, Latvia, and Ukraine), but later naturalized and started to spread into anthropogenic, semi-natural, and natural habitats [37,42,43,53,54]. We presume that, in Lithuania, subsp. australis had a lag phase of 40–50 years [60], with intensive spread starting in the 1990s. Initially, the plants spread close to their growing sites, whereas intensive cryptic spread over significant distances from the original stands started about two decades ago.
The current wide distribution of subsp. australis in Lithuania is primarily the result of its long history of large-scale planting along railways, roads, and ornamental plantations [37]. This has led to many seed sources, which in turn have led to high propagule pressure [61]. The most important vector of dispersal is frugivorous birds which, by feeding on fleshy fruits, disperse seeds not only around the feeding grounds but also over much greater distances [62,63]. Plant waste from gardens, which includes not only fallen leaves but also the seeds and fruits of cultivated plants, is sometimes dumped at the edges of forests, in shrublands, along roads, in forest clearings, or near water bodies. As the plant debris rots, some of the seeds germinate and form new dense stands of subsp. australis. During our investigations, we found several stands of such origin at significant distances from known sites of cultivation (e.g., in Kaišiadorys, Kaunas, and Kupiškis districts).
In Lithuania, we did not find mixed stands of individuals of both subspecies of Cornus sanguinea. If the alien subsp. australis continues to spread, the formation of mixed stands is likely to occur, which could lead to interbreeding with the native subsp. sanguinea. In this case, there would be a threat, as hybrid populations might respond differently to ecological conditions (Pyšek et al., 2013). Some researchers consider that such hybrids already exist in Central Europe and are called Cornus sanguinea subsp. hungarica (Kárpáti) Soó [45,64]. There is no strong support for the hybrid origin of subsp. hungarica yet, but it is assumed that the plants are representatives of the polymorphic subsp. sanguinea [42]. Nevertheless, the consequences of possible hybridization between native and alien subspecies are difficult to predict.
The current area of occurrence of subsp. australis covers about 63% of the territory of Lithuania, and there are only a few areas of the country where this subspecies has not been detected. The current distribution, invaded areas, and high plant abundance in some habitats suggest that subsp. australis will continue to spread in Lithuania. Some nurseries in the country still sell saplings of these plants, which are mainly used for hedgerow formation. This further increases the number of introduction sites and thus propagule pressure. Since the seeds of Cornus species are efficiently dispersed by frugivorous birds [62,63,65], an even more rapid spread and invasion of subsp. australis into new areas and habitats can be anticipated.
4.2. Comparison of Reproductive Traits of Subspecies
4.2.1. Flowering and Fruiting
Propagule pressure and the actual fecundity of individuals are particularly important determinants of invasiveness [22,61]. Many alien species tend to perform better in an invasive range than in their native range, or their performance in an invasive range is not substantially altered [66,67,68]. There are no data available regarding the potential and realized fecundity of subsp. australis in its native range; therefore, we can only compare these characteristics with those of the native subsp. sanguinea.
The results of this study showed that alien subsp. australis inflorescences in Lithuania have significantly more flowers than those of the native subsp. sanguinea. A comparison of the results of this study with analogous data from Spain [69] showed that there were no significant differences between regions in terms of the mean number of flowers per inflorescence (t = 1.78; p = 0.08) of subsp. sanguinea. The number of flowers per inflorescence of subsp. australis was also significantly higher in Lithuania compared with subsp. sanguinea from Spain (t = 31.87; p < 0.001). Since the mean number of flowers of subsp. sanguinea was similar in Lithuania and Spain, and the mean number of flowers per inflorescence was significantly lower than the mean number of flowers of subsp. australis, it can be assumed that different numbers of flowers per inflorescence is a characteristic of the subspecies. However, this assumption could only be confirmed or rejected by a much larger study including populations of subsp. australis from its native range.
An analysis of the results of the number of fruits per infructescence showed that, in Lithuania, the number of fruits was significantly higher for subsp. australis than subsp. sanguinea. A comparison of the results of our study with the results of an analogous study in Spain [69] (Guitián et al., 1996) showed that the mean number of fruits per infructescence was lower than that in Lithuania for both subsp. sanguinea (t = 18.22; p < 0.001) and subsp. australis (t = 31.87; p < 0.001). However, in Lithuania, the subsp. sanguinea was found to have a higher realized fecundity (36.6% of the potential fecundity) than that of subsp. australis (30.5%), and both were higher than the realized fecundity of subsp. sanguinea in Spain (21.8% of the potential fecundity). These differences in realized fecundity in Lithuania and Spain could have various causes. It is known that realized fecundity depends on the individual characteristics of the plant, the habitat conditions, the meteorological conditions of the flowering period, the fruit ripening period, the abundance of pollinators in the area, etc. [68,70].
Although the realized fecundity of native subsp. sanguinea was higher than that of the alien subsp. australis, the real number of ripened fruits per subsp. australis infructescence was significantly higher. This results in higher pressure of propagules for subsp. australis than subsp. sanguinea, which is directly related to the rate of spread [22]. The high propagule pressure of subsp. australis has been confirmed by studies on the composition of the soil seed bank of Cornus species performed in Lithuania [71]. Thus, the results of the study on flowering and fruit production confirm that the alien subsp. australis has an advantage over the native subsp. sanguinea.
4.2.2. Age of Maturity of Subspecies
A short juvenile period of individuals of alien species, especially woody plants, is one of the characteristics that determine their invasiveness [14]. Comparing the age at which individuals of the two subspecies of Cornus sanguinea reach the generative stage, we found that subsp. australis individuals start flowering and fruiting at the age of three years, while the youngest individuals of subsp. sanguinea reach this stage at the age of four years. Although the juvenile age of both species is short, subsp. australis begins fruiting at one year younger than subsp. sanguinea growing in open habitats. Given that the native subsp. sanguinea usually grows in forests [61], while the alien subsp. australis spreads primarily into open habitats [53,72], the difference in age of maturity between the two subspecies may be even greater. In addition, to reach generative maturity, the native subsp. sanguinea needs to invest more resources for growth [73] and reach a larger stem diameter than subsp. australis.
A study of the demographic composition of populations of the invasive shrub Cytisus scoparius (L.) Link in Lithuania has shown that the youngest individuals to reach the generative stage were three years old [16]. In the Pampas region of Argentina, the age of individuals of the invasive Prunus mahaleb L. has been studied, and it was found that the youngest plants producing their first fruits were also three years old [74]. The lifespan and reproductive age of woody plants are known to vary from a few to several decades, depending on the species. Thus, a short juvenile period further extends the duration of an individual’s reproductive stage, and newly established populations increase propagule pressure and the potential for further spread in a shorter time, as well as the ability of the population to recover after natural death or the targeted elimination of adult individuals. The early maturation of young woody plant individuals has a significant impact on the choice of control or eradication measures for an invasive species [75].
The results of this study show that the dendrochronological method can be easily applied to assess the development and maturation of woody alien species. The growth and maturation of individuals can also be assessed under experimental (ex situ) conditions, but more accurate information on the characteristics of the plant can be obtained when it is assessed under natural (in situ) conditions [76,77]. In addition, this method allows us to assess not only the age of individuals and the demographic composition of populations [16] but also the growth rate of individuals under different habitat conditions and their response to changes in meteorological or other environmental conditions [15,74].
4.3. Species Diversity in Invaded Communities
Some studies have shown that native dominant species have the same negative impact on the species diversity of communities as dominant alien species [30]. As we studied two subspecies of the same species, it might have been expected that their effects on species diversity would be the same under similar or identical environmental conditions. In this study, plant communities with alien subsp. australis were similar, in most characteristics, to native subsp. sanguinea. The only significant difference was that some of the communities were either abundant or dominated by alien subsp. australis, while others had a similar abundance of native subsp. sanguinea. Herbaceous plant species diversity was significantly lower in communities with subsp. australis, and there was a significant negative correlation between the abundance of subsp. australis and species diversity, while the abundance of subsp. sanguinea had no significant effect on species diversity in the communities. We assume that subsp. australis may have a stronger negative effect on species diversity due to its stronger effect on soil humidity and the availability of nutrients than subsp. sanguinea [78]. However, the diversity of woody plant species was significantly higher in communities with subsp. australis than in communities with subsp. sanguinea and the negative impact on herbaceous species diversity may be caused by the cumulative effect of all woody plants [79].
The number of alien species in the communities studied with subsp. australis was significantly higher than in the plots with subsp. sanguinea, but the overall frequency of alien species was low in all communities studied. A higher proportion of alien species in the communities with subsp. australis may be explained by the fact that they were surveyed closer to urbanized areas, where the number of alien species is always higher, compared with the remote areas [80] where most of the communities with subsp. sanguinea were studied. Based on the results of this study, we cannot identify any clear association between subsp. australis and other alien species.
5. Conclusions
In summary, all available information has revealed that the alien Cornus sanguinea subsp. australis, which has been planted in shelter and ornamental plantations, has spread unnoticed throughout most of Lithuania for several decades. The cryptic invasion has been influenced by a large number of introduction sites, the short juvenile period of the subspecies, and its high production of seeds, which are mostly dispersed by birds.
An evaluation of reproductive traits confirmed that the root suckers of subsp. australis reach generative maturity at a younger age, and with a smaller diameter of root suckers, than Cornus sanguinea subsp. sanguinea. Furthermore, the inflorescences of subsp. australis had significantly more flowers than the inflorescences of subsp. sanguinea and ripened significantly more fruit in the infructescences.
The results of the study also demonstrate the importance of using the seeds of trees and shrubs from local provenance for the establishment of various types of plantations and afforestation. Even when seeds or seedlings of native tree and shrub species are introduced from other regions, a high risk of the cryptic invasion of alien subspecies, races, and genotypes remains.
Author Contributions
Conceptualization, L.P. and Z.G.; methodology, L.P. and Z.G.; formal analysis, L.P. and Z.G.; investigation, L.P. and Z.G.; data curation, L.P.; writing—original draft preparation, L.P. and Z.G.; writing—review and editing, L.P. and Z.G.; visualization, L.P. and Z.G.; supervision, Z.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Richardson, D.M. Pinus: A model group for unlocking the secrets of alien plant invasions? Preslia 2006, 78, 375–388. [Google Scholar]
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Follak, S.; Bakacsy, L.; Essl, F.; Hochfellner, L.; Lapin, K.; Schwarz, M.; Tokarska-Guzik, B.; Wołkowycki, D. Monograph of invasive plants in Europe N°6: Asclepias syriaca L. Bot. Lett. 2021, 168, 422–451. [Google Scholar] [CrossRef]
- Verloove, F. Invaders in disguise. Conservation risks derived from misidentifications of invasive plants. Manag. Biol. Invasions 2010, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Saltonstall, K. Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. PNAS 2002, 99, 2445–2449. [Google Scholar] [CrossRef] [Green Version]
- Morais, P.; Reichard, M. Cryptic invasions: A review. Sci. Total Environ. 2018, 613, 1438–1448. [Google Scholar] [CrossRef]
- Jarić, I.; Heger, T.; Monzon, F.C.; Jeschke, J.M.; Kowarik, I.; McConke, K.R.; Pyšek, P.; Sagouis, A.; Essl, F. Crypticity in biological invasions. Trends Ecol. Evol. 2019, 34, 291–302. [Google Scholar] [CrossRef]
- Canavan, K.; Canavan, S.; Harms, N.E.; Lambertini, C.; Paterson, I.D.; Thum, R. The potential for biological control on cryptic plant invasions. Biol. Control 2020, 144, 104243. [Google Scholar] [CrossRef]
- Wahlsteen, E. Morphometrical characteristics of cryptic invasive and indigenous gene pools of field maple Acer campestre L. in southern Sweden. Nord. J. Bot. 2021, 39, e02901. [Google Scholar] [CrossRef]
- Hyldgaard, B.; Lambertini, C.; Brix, H. Phylogeography reveals a potential cryptic invasion in the Southern Hemisphere of Ceratophyllum demersum, New Zealand’s worst invasive macrophyte. Sci. Rep. 2017, 7, 16569. [Google Scholar] [CrossRef] [Green Version]
- Boeraeve, M.; D’hondt, B.; Verloove, F.; Vandekerkhove, K.; Van Landuyt, W.; Tamsyn, W.; Adriaens, T. “Lianification” or liana invasion? Commentary on Perring et al. (2020). Front. Ecol. Environ. 2021, 19, 375–376. [Google Scholar] [CrossRef]
- van Kleunen, M.; Weber, E.; Fischer, M. A meta analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, S.I.; Richardson, D.M. Invasive plants have broader physiological niches. PNAS 2014, 111, 10610–10614. [Google Scholar] [CrossRef] [Green Version]
- Rejmánek, M.; Richardson, D.M. What attributes make some plant species more invasive? Ecology 1996, 77, 1655–1661. [Google Scholar] [CrossRef]
- Gudžinskas, Z.; Petrulaitis, L.; Žalneravičius, E. Emerging invasion threat of the liana Celastrus orbiculatus (Celastraceae) in Europe. NeoBiota 2020, 5, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Taura, L.; Gudžinskas, Z. Life stages and demography of invasive shrub Cytisus scoparius (Fabaceae) in Lithuania. Botanica 2020, 26, 1–14. [Google Scholar] [CrossRef]
- Xavier, C.N.; Granato-Souza, D.; Barbosa, A.C.; da Silva, J.R.M. Tropical dendrochronology applied to invasive tree species in the Brazilian Atlantic Forest. J. For. Res. 2021, 32, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Silveri, A.; Dunwiddie, P.W.; Michaels, H.J. Logging and edaphic factors in the invasion of an Asian woody vine in a mesic North American forest. Biol. Invasions 2001, 3, 379–389. [Google Scholar] [CrossRef]
- Mestre, L.; Chartier, M.; Renison, D. Effect of livestock on the growth of Rosa rubiginosa in a mountain range: A dendrochronological approach. Appl. Ecol. Environ. Res. 2014, 12, 855–866. [Google Scholar] [CrossRef]
- Richardson, D.M. Forestry trees as invasive aliens. Conserv. Biol. 1998, 12, 18–26. [Google Scholar] [CrossRef]
- van Kleunen, M.; Essl, F.; Pergl, J.; Brundu, G.; Carboni, M.; Dullinger, S.; Early, R.; González-Moreno, P.; Groom, Q.J.; Hulme, P.E.; et al. The changing role of ornamental horticulture in alien plant invasions. Biol. Rev. 2018, 93, 1421–1437. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Křivánek, M.; Jarošík, V. Planting intensity, residence time, and species traits determine invasion success of alien woody species. Ecology 2009, 90, 2734–2744. [Google Scholar] [CrossRef] [PubMed]
- Collete, L.K.D.; Pither, J. Russian-olive (Elaeagnus angustifolia) Biology and ecology and its potential to invade Northern North American riparian ecosystems. Invasive Plant Sci. Manag. 2015, 8, 1–14. [Google Scholar] [CrossRef]
- Deckers, B.; Verheyen, K.; Hermy, M.; Muys, B. Effects of landscape structure on the invasive spread of black cherry Prunus serotina in an agricultural landscape in Flanders, Belgium. Ecography 2005, 28, 99–109. [Google Scholar] [CrossRef]
- Le Viol, I.; Julliard, R.; Kerbiriou, C.; De Redon, L.; Carnino, N.; Machon, N.; Porcher, E. Plant and spider communities benefit differently from the presence of planted hedgerows in highway verges. Biol. Conserv. 2008, 141, 1581–1590. [Google Scholar] [CrossRef] [Green Version]
- Blanusa, T.; Garratt, M.; Cathcart-James, M.; Hunt, L.; Cameron, R.W. Urban hedges: A review of plant species and cultivars for ecosystem service delivery in north-west Europe. Urban For. Urban Green 2019, 44, 126391. [Google Scholar] [CrossRef]
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D.; et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.L.; Standish, R.J.; Stock, W.D.; Valladares, F. Plant functional traits of dominant native and invasive species in mediterranean-climate ecosystems. Ecology 2016, 97, 75–83. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Hejda, M.; Sádlo, J.; Kutlvašr, J.; Petřík, P.; Vítková, M.; Vojík, M.; Pyšek, P.; Pergl, J. Impact of invasive and native dominants on species richness and diversity of plant communities. Preslia 2021, 93, 181–201. [Google Scholar] [CrossRef]
- Tanner, R.; Branquart, E.; Brundu, G.; Buholzer, S.; Chapman, D.; Ehret, P.; van Valkenburg, J. The prioritisation of a short list of alien plants for risk analysis within the framework of the Regulation (EU) No. 1143/2014. NeoBiota 2017, 35, 87–118. [Google Scholar] [CrossRef]
- Yazlık, A.; Ambarlı, D. Do non-native and dominant native species carry a similar risk of invasiveness? A case study for plants in Turkey. NeoBiota 2022, 76, 53–72. [Google Scholar] [CrossRef]
- Kowarik, I. Time lags in biological invasions with regard to the success and failure of alien species. In Plant Invasions: General Aspects and Special Problems; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 15–38. [Google Scholar]
- Petrulaitis, L.; Gudžinskas, Z. The first records of two alien woody species, Cornus alternifolia and Cornus amomum, in Lithuania. BioInv Rec. 2020, 9, 384–392. [Google Scholar] [CrossRef]
- Medvecká, J.; Kliment, J.; Májeková, J.; Halada, Ľ.; Zaliberová, M.; Gojdičová, E.; Feráková, V.; Jarolímek, I. Inventory of the alien flora of Slovakia. Preslia 2012, 84, 257–309. [Google Scholar]
- Azola, T.; Bakan, B. Popis tujerodnih rastlinskih vrst v Mestni občini Maribor [Inventory of alien plant species in the Municipality of Maribor]. Hladnikia 2021, 47, 3–16. [Google Scholar]
- Gudžinskas, Z.; Petrulaitis, L. New alien taxa of the genus Cornus (Cornaceae) recorded in Lithuania and Latvia. Botanica 2021, 27, 160–169. [Google Scholar] [CrossRef]
- Kelly, D.L. Cornus sericea L. in Ireland: An incipient weed of wetlands. Watsonia 1990, 18, 33–36. [Google Scholar]
- Bačič, T.; Strgulc Krajšek, S.; Jogan, N. Red osier dogwood (Cornus sericea L.)—A new invasive species in Slovenian flora. Acta Biol. Slov. 2015, 58, 13–21. (In Slovenian) [Google Scholar]
- Bomanowska, A.; Adamowski, W.; Kirpluk, I.; Otreba, A.; Rewicz, A. Invasive alien plants in Polish national parks—Threats to species diversity. PeerJ 2019, 7, e8034. [Google Scholar] [CrossRef] [Green Version]
- Gregor, T.; Kasperek, G. Non-native woody plant species in urban forests of Frankfurt/Main (Germany). Tuexenia 2021, 41, 133–145. [Google Scholar] [CrossRef]
- Riebl, R.; Meve, U.; Aas, G. Morphologische Variabilität und taxonomische Differenzierung von Cornus sanguinea: Nordbayerische Naturstandorte und Strassenbegleitgrün im Vergleich. Ber. Bayer. Bot. Ges. Erforsch. Heim. Flora 2017, 87, 39–54. [Google Scholar]
- Verloove, F. Fraxinus pennsylvanica, Pterocarya fraxinifolia en andere opmerkelijke uitheemse rivierbegeleiders in België en NW-Frankrijk. Dumortiera 2011, 99, 1–10. [Google Scholar]
- Poyarkova, A.I. Cornaceae. In Flora of USSR; Shishkin, B.K., Ed.; Smithsonian Libraries: Washington, DC, USA, 1973; Volume 17, pp. 225–249. [Google Scholar]
- Eyde, R.H. Comprehending Cornus: Puzzles and progress in the systematics of dogwoods. Bot. Rev. 1988, 54, 233–351. [Google Scholar] [CrossRef]
- Woźnicka, A.; Melosik, I.; Morozowska, M. Quantitative and qualitative differences in morphological traits of endocarps revealed between Cornus L. species. Plant Syst. Evol. 2015, 301, 291–308. [Google Scholar] [CrossRef] [Green Version]
- Morozowska, M.; Woźnicka, A.; Nowińska, R. The morphology and internal structure of dogwood (Cornus L.) endocarps in the taxonomy and phylogeny of the genus. PeerJ 2021, 9, e12170. [Google Scholar] [CrossRef]
- Schulz, B. Die Gattung Cornus (Cornaceae), Hartriegel und Kornelkirsche, Teil 3. Mitt. Deutsch. Dendrol. Ges. 2012, 97, 91–132. [Google Scholar]
- Grosset, E.G. The geographical distribution of Cornus sanguinea s. l. and history of the broad-leaved forests of the European part of the U.S.S.R. Bull. Soc. Nat. Mosc. Biol. Ser. 1958, 63, 77–85. [Google Scholar]
- Novák, P.; Zukal, D.; Harásek, M.; Vlčková, P.; Abdaladze, O.; Willner, W. Ecology and vegetation types of oak-hornbeam and ravine forests of the Eastern Greater Caucasus, Georgia. Folia Geobot. 2020, 55, 333–349. [Google Scholar] [CrossRef]
- Essl, F.; Bacher, S.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Katsanevakis, S.; Kowarik, I.; Kühn, I.; Pyšek, P.; Rabitsch, W.; et al. Which taxa are alien? Criteria, applications, and uncertainties. Bioscience 2018, 68, 496–509. [Google Scholar] [CrossRef] [Green Version]
- Maes, B.; van Loon, R. Wilde en niet-wilde bomen en struiken in vegetatieopnamen. Een voorstel voor een methode van specifieke notering van bomen en struiken in vegetatieopnamen en overwegingen daarbij. Stratiotes 2020, 55, 19–31. [Google Scholar]
- Shynder, O.; Negrash, J. Checklist of the flora of the vicinity of Balakliya (Kharkiv region, Ukraine): Native and alien taxa, distribution of rare plants, new findings. Plant Introd. 2021, 89–90, 13–71. [Google Scholar] [CrossRef]
- Orlov, O.O.; Shynder, O.I.; Vorobjov, E.O.; Gryb, O.V. New floristic finds in the Forest-Steppe part of Zhytomyr Region. Ukr. Bot. J. 2022, 79, 6–26. [Google Scholar] [CrossRef]
- Convention on Biological Diversity. Pathways of Introduction of Invasive Species, Their Prioritization and Management. 2014. Available online: https://www.cbd.int/doc/meetings/sbstta/sbstta-18/official/sbstta-18-09-add1-en.pdf (accessed on 25 October 2022).
- Gudžinskas, Z. Genus Ambrosia L. (Asteraceae) in Lithuania. Thaiszia 1993, 3, 89–96. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie: Grundzüge der Vegetationskunde. Dritte Auflage; Springer: New York, NY, USA, 1964; p. 865. [Google Scholar] [CrossRef]
- Lenth, R.V. Some practical guidelines for effective sample size determination. Am. Stat. 2001, 55, 187–193. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Crooks, J. Lag times and exotic species: The ecology and management of biological invasions in slow-motion. Ecoscience 2005, 12, 316–329. [Google Scholar] [CrossRef]
- Simberloff, D. The role of propagule pressure in biological invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Krüsi, B.O.; Debussche, M. The fate of flowers and fruits of Cornus sanguinea L. in three contrasting Mediterranean habitats. Oecologia 1988, 74, 592–599. [Google Scholar] [CrossRef]
- Kollmann, J.; Grubb, P.J. Biological flora of Central Europe: Cornus sanguinea L. Flora 2001, 196, 161–179. [Google Scholar] [CrossRef]
- Holub, J. Cornaceae Dumort.—Dřínovité. In Květena České republiky; Slavík, B., Chrtek, J., Tomšovic, P., Eds.; Academia: Praha, Czech Republic, 1997; Volume 5, pp. 252–264. [Google Scholar]
- Borowicz, V.A.; Stephenson, A.G. Fruit composition and patterns of fruit dispersal of two Cornus spp. Oecologia 1985, 67, 435–441. [Google Scholar] [CrossRef]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Parker, J.D.; Torchin, M.E.; Hufbauer, R.A.; Lemoine, N.P.; Alba, C.; Blumenthal, D.M.; Bossdorf, O.; Byers, J.E.; Dunn, A.M.; Heckman, R.W.; et al. Do invasive species perform better in their new ranges? Ecology 2013, 94, 985–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudžinskas, Z.; Taura, L. Do Reproductive Traits of Invasive Populations of Scotch Broom, Cytisus scoparius (Fabaceae), Outperform Native Populations? Plants 2022, 11, 2158. [Google Scholar] [CrossRef] [PubMed]
- Guitián, J.; Guitián, P.; Navarro, L. Fruit set, fruit reduction, and fruiting strategy in Cornus sanguinea (Cornaceae). Am. J. Bot. 1996, 83, 744–748. [Google Scholar] [CrossRef]
- Schurr, F.M.; Steinitz, O.; Nathan, R. Plant fecundity and seed dispersal in spatially heterogeneous environments: Models, mechanisms and estimation. J. Ecol. 2008, 96, 628–641. [Google Scholar] [CrossRef]
- Petrulaitis, L.; Rašomavičius, V.; Uogintas, D.; Gudžinskas, Z. Soil seed bank of alien and native Cornus (Cornaceae) taxa in Lithuania: What determines seed density and vertical distribution in soil? Diversity 2022, 14, 488. [Google Scholar] [CrossRef]
- Verloove, F. Cornus sanguinea. From: Manual of the Alien Plants of Belgium. Botanic Garden Meise, Belgium. 2021. Available online: www.alienplantsbelgium.be (accessed on 15 October 2021).
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand? In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 193, pp. 97–125. [Google Scholar] [CrossRef]
- Zalba, S.M.; Amodeo, M.R. Site-specific reproductive performance of an invasive fleshy-fruited tree in remnant grasslands of the Argentinean Pampas. Weed Res. 2015, 55, 546–554. [Google Scholar] [CrossRef]
- Serviss, B.E.; Tumlison, R. Guide to the naturalized, escaped, and adventive woody flora of Arkansas. Phytoneuron 2021, 29, 1–193. [Google Scholar]
- Grubb, P.J.; Kollmann, J.; Lee, W.G. A Garden Experiment on Susceptibility to Rabbit-Grazing, Sapling Growth Rates, and Age at First Reproduction for Eleven European Woody Species. Plant Biol. 1999, 1, 226–234. [Google Scholar] [CrossRef]
- Chevin, L.-M.; Collins, S.; Lefèvre, F. Phenotypic plasticity and evolutionary demographic responses to climate change: Taking theory out to the field. Funct. Ecol. 2013, 27, 967–979. [Google Scholar] [CrossRef] [Green Version]
- He, Y. The Relationship between an Invasive Shrub and Soil Moisture: Seasonal Interactions and Spatially Covarying Relations. ISPRS Int. J. Geo-Inf. 2014, 3, 1139–1153. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Classen, A.T.; Simberloff, D. Two co-occurring invasive woody shrubs alter soil properties and promote subdominant invasive species. J. Appl. Ecol. 2014, 51, 124–133. [Google Scholar] [CrossRef] [Green Version]
- De Barros Ruas, R.; Costa, L.M.S.; Bered, F. Urbanization driving changes in plant species and communities—A global view. Glob. Ecol. Conserv. 2022, 38, e02243. [Google Scholar] [CrossRef]
Figure 1.
Position of Lithuania in Europe (left) and location of the Cornus sanguinea subsp. sanguinea (empty circles) and Cornus sanguinea subsp. australis (black dots) sampling sites for reproductive traits in Lithuania (right).
Figure 1.
Position of Lithuania in Europe (left) and location of the Cornus sanguinea subsp. sanguinea (empty circles) and Cornus sanguinea subsp. australis (black dots) sampling sites for reproductive traits in Lithuania (right).

Figure 2.
The current distribution of Cornus sanguinea subsp. australis in Lithuania. The red dot marks the first record of this taxon in Lithuania; the black dots mark localities based on the results of this research.
Figure 2.
The current distribution of Cornus sanguinea subsp. australis in Lithuania. The red dot marks the first record of this taxon in Lithuania; the black dots mark localities based on the results of this research.

Figure 3.
Comparison of the number of flowers (a) and the number of fruits (b) at the study sites of Cornus sanguinea subsp. sanguinea (A—Piepaliai; B—Veliuona; C—Šilinė) and Cornus sanguinea subsp. australis (D—Maišiagala; E—Ginkūnai; F—Kaišiadorys). Different lower-case letters denote statistically significant differences between the means among the sites, applying the Mann–Whitney U-test.
Figure 3.
Comparison of the number of flowers (a) and the number of fruits (b) at the study sites of Cornus sanguinea subsp. sanguinea (A—Piepaliai; B—Veliuona; C—Šilinė) and Cornus sanguinea subsp. australis (D—Maišiagala; E—Ginkūnai; F—Kaišiadorys). Different lower-case letters denote statistically significant differences between the means among the sites, applying the Mann–Whitney U-test.

Figure 4.
Comparison of the number of herb species (a) and the number of alien species (b) between study plots with Cornus sanguinea subsp. australis and Cornus sanguinea subsp. sanguinea. Asterisks above the whiskers indicate the differences between all study plots according to the results of the two-sample Mann–Whitney test (*—p < 0.05; ***—p < 0.001).
Figure 4.
Comparison of the number of herb species (a) and the number of alien species (b) between study plots with Cornus sanguinea subsp. australis and Cornus sanguinea subsp. sanguinea. Asterisks above the whiskers indicate the differences between all study plots according to the results of the two-sample Mann–Whitney test (*—p < 0.05; ***—p < 0.001).

Figure 5.
Relationship between the coverage of Cornus sanguinea subsp. australis (a) and Cornus sanguinea subsp. sanguinea (b) and the number of herbaceous plant species per plot.
Figure 5.
Relationship between the coverage of Cornus sanguinea subsp. australis (a) and Cornus sanguinea subsp. sanguinea (b) and the number of herbaceous plant species per plot.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Locations of Cornus sanguinea subsp. sanguinea and Cornus sanguinea subsp. australis sampling for the study of reproductive traits in Lithuania.
Table 1.
Locations of Cornus sanguinea subsp. sanguinea and Cornus sanguinea subsp. australis sampling for the study of reproductive traits in Lithuania.
| Subspecies | Site Name | Administrative Unit | Longitude (°E) | Latitude (°N) |
|---|---|---|---|---|
| Cornus sanguinea subsp. sanguinea | Piepaliai | Kaunas distr. | 55.07423 | 23.78346 |
| Cornus sanguinea subsp. sanguinea | Veliuona | Jurbarkas distr. | 55.08996 | 23.18326 |
| Cornus sanguinea subsp. sanguinea | Šilinė | Jurbarkas distr. | 55.08856 | 22.94780 |
| Cornus sanguinea subsp. australis | Ginkūnai | Šiauliai distr. | 55.95343 | 23.37109 |
| Cornus sanguinea subsp. australis | Kaišiadorys | Kaišiadorys city | 54.87099 | 24.43677 |
| Cornus sanguinea subsp. australis | Maišiagala | Vilnius distr. | 54.86959 | 25.05138 |
Table 2.
Comparison of the age of the youngest fruiting shoots and the diameter of Cornus sanguinea subsp. australis and Cornus sanguinea subsp. sanguinea at the study sites. Different lowercase superscript letters indicate significant differences in the mean values between sites and different capital letters between subspecies relate to the results of the Mann–Whitney U-test.
Table 2.
Comparison of the age of the youngest fruiting shoots and the diameter of Cornus sanguinea subsp. australis and Cornus sanguinea subsp. sanguinea at the study sites. Different lowercase superscript letters indicate significant differences in the mean values between sites and different capital letters between subspecies relate to the results of the Mann–Whitney U-test.
| Taxa and Sites | Age (years) | Shoot Diameter (mm) | ||||
|---|---|---|---|---|---|---|
| Min–Max | Mean | Median | Min–Max | Mean | Median | |
| Cornus sanguinea subsp. australis | ||||||
| Kaišiadorys | 3–6 | 3.9 ± 0.7 a | 4 | 5.0–8.8 | 7.18 ± 1.28 a | 7.7 |
| Vievis | 3–5 | 3.5 ± 0.6 a | 3 | 4.9–8.8 | 6.01 ± 0.90 b | 5.7 |
| Pooled | 3–6 | 3.7 ± 0.7 A | 4 | 4.9–8.8 | 6.60 ± 1.24 A | 6.5 |
| Cornus sanguinea subsp. sanguinea | ||||||
| Piepaliai | 4–6 | 5.4 ± 0.7 b | 5 | 7.5–13.0 | 10.90 ± 1.62 c | 11.3 |
| Veliuona | 4–6 | 5.2 ± 0.6 b | 5 | 8.0–14.6 | 12.18 ± 1.79 d | 12.5 |
| Pooled | 4–6 | 5.3 ± 0.1 B | 5 | 7.5–14.6 | 11.55 ± 1.80 B | 11.9 |
Table 3.
Results of a two-way nested ANOVA analysis showing the effect of the taxon (fixed effect: f) and habitat (random effect: r) on the age and diameter of the shoots that had started fruiting.
Table 3.
Results of a two-way nested ANOVA analysis showing the effect of the taxon (fixed effect: f) and habitat (random effect: r) on the age and diameter of the shoots that had started fruiting.
| Factor | Sum of Squares | df | Mean Square | F | p |
|---|---|---|---|---|---|
| Shoot age | |||||
| Taxon (f) | 76.80 | 1 | 76.80 | 168.30 | <0.001 |
| Habitat (r) | 2.26 | 2 | 1.13 | 2.48 | 0.088 |
| Within | 52.93 | 116 | 0.46 | ||
| Total | 132.00 | 119 | |||
| Shoot diameter | |||||
| Taxon (f) | 737.55 | 1 | 737.55 | 357.00 | <0.001 |
| Habitat (r) | 43.84 | 2 | 21.92 | 10.61 | <0.001 |
| Within | 239.67 | 116 | 2.07 | ||
| Total | 1021.06 | 119 |
Table 4.
Statistical comparison of the characteristics (mean ± standard deviation) of the plant communities in the study plots with Cornus sanguinea subsp. australis and Cornus sanguinea subsp. sanguinea. Differences between all study plots were determined with a two-sample Mann–Whitney test (*—p < 0.05; ***—p < 0.001; ns—not significant). Comparisons between habitat groups were performed applying the Mann–Whitney pairwise comparison and different letters indicate significant differences between the two samples.
Table 4.
Statistical comparison of the characteristics (mean ± standard deviation) of the plant communities in the study plots with Cornus sanguinea subsp. australis and Cornus sanguinea subsp. sanguinea. Differences between all study plots were determined with a two-sample Mann–Whitney test (*—p < 0.05; ***—p < 0.001; ns—not significant). Comparisons between habitat groups were performed applying the Mann–Whitney pairwise comparison and different letters indicate significant differences between the two samples.
| Parameters | All Plots | Woodlands | Shrublands | Woodlands | Shrublands | |
|---|---|---|---|---|---|---|
| Cornus sanguinea | Cornus sanguinea | Cornus sanguinea | ||||
| subsp. australis | subsp. sanguinea | subsp. australis | subsp. australis | subsp. sanguinea | subsp. sanguinea | |
| Cover of Cornus (%) | 56.6 ± 21.6 ns | 63.5 ± 23.5 ns | 59.2 ± 21.2 a | 52.3 ± 22.6 a | 65.9 ± 24.2 a | 60.7 ± 23.0 a |
| Cover of trees (%) | 37.2 ± 25.7 ns | 31.6 ± 30.7 ns | 53.7 ± 14.9 a | 8.6 ± 10.0 b | 57.3 ± 14.5 a | 0.4 ± 1.3 c |
| Cover of shrubs (%) | 64.7 ± 24.9 * | 78.1 ± 17.1 * | 65.8 ± 26.1 a | 62.7 ± 23.7 a | 76.5 ± 19.0 a | 80.0 ± 14.8 a |
| Cover of herbaceous species (%) | 38.5 ± 31.8 ns | 42.1 ± 28.0 ns | 32.0 ± 29.1 a | 49.6 ± 34.4 ab | 31.7 ± 22.9 a | 54.6 ± 29.2 b |
| Cover of bryophytes (%) | 13.7 ± 20.3 ns | 26.1 ± 30.8 ns | 13.2 ± 17.9 a | 14.6 ± 24.9 a | 22.2 ± 27.9 a | 30.8 ± 34.4 a |
| Total number of plant species | 22.6 ± 8.2 ns | 24.6 ± 6.4 ns | 22.9 ± 7.6 a | 22.2 ± 9.5 a | 23.4 ± 7.1 a | 26.1 ± 5.3 a |
| Number of native vascular plant species | 15.4 ± 8.8 *** | 22.7 ± 6.2 *** | 15.9 ± 6.4 ns | 14.4 ±12.1 ns | 21.5 ± 6.7 ns | 24.1 ± 5.3 ns |
| Total number of woody species | 8.2 ± 4.3 ns | 7.0 ± 2.7 ns | 9.9 ± 4.1 a | 5.3 ± 2.9 b | 8.3 ± 2.5 a | 5.4 ± 2.0 b |
| Total number of herbaceous plant species | 12.9 ± 6.8 * | 15.8 ± 5.8 * | 11.6 ± 5.0 a | 15.3 ± 8.9 ab | 13.0 ± 4.7 a | 19.2 ± 5.3 b |
| Number of alien species | 3.5 ± 1.5 *** | 0.6 ± 1.0 *** | 3.8 ± 1.6 a | 2.9 ± 1.4 a | 0.5 ± 0.9 b | 0.8 ± 1.2 b |
| Number of alien woody species | 3.1 ± 1.6 *** | 0.3 ± 0.5 *** | 3.6 ± 1.6 a | 2.3 ± 1.0 b | 0.2 ± 0.4 c | 0.4 ± 0.6 c |
| Number of alien herbaceous species | 0.4 ± 0.7 ns | 0.4 ± 0.6 ns | 0.3 ± 0.4 a | 0.6 ± 1.0 a | 0.3 ± 0.6 a | 0.4 ± 0.8 a |
| Number of bryophyte species | 1.5 ± 1.5 ns | 1.2 ± 1.6 ns | 1.4 ± 1.5 a | 1.6 ± 1.6 a | 1.6 ± 1.6 a | 0.7 ± 1.3 a |
Table 5.
The number of plant species and their frequency (%) in the study plots of plant communities with Cornus sanguinea subsp. australis and Cornus sanguinea subsp. sanguinea. The table includes species with a frequency of more than 40% in at least one group of the study plots.
Table 5.
The number of plant species and their frequency (%) in the study plots of plant communities with Cornus sanguinea subsp. australis and Cornus sanguinea subsp. sanguinea. The table includes species with a frequency of more than 40% in at least one group of the study plots.
| Group of Habitats | Woodlands | Shrublands | ||
|---|---|---|---|---|
| Subspecies | Cornus sanguinea | Cornus sanguinea | ||
| subsp. australis | subsp. sanguinea | subsp. australis | subsp. sanguinea | |
| Number of study plots | 19 | 16 | 11 | 14 |
| Total number of species | 160 | 136 | 140 | 170 |
| Total number of woody species | 55 | 33 | 29 | 30 |
| Total number of herbaceous species | 94 | 95 | 90 | 132 |
| Number of bryophyte species | 11 | 8 | 11 | 8 |
| Total number of alien species | 27 | 6 | 15 | 9 |
| Number of alien woody species | 25 | 3 | 10 | 4 |
| Number of alien herbaceous species | 2 | 3 | 5 | 5 |
| Woody species | ||||
| Cornus sanguinea L. subsp. australis (C.A.Mey.) Jáv. | 100 | 0 | 100 | 0 |
| Cornus sanguinea L. subsp. sanguinea | 0 | 100 | 0 | 100 |
| Acer platanoides L. | 90 | 65 | 27 | 21 |
| Rubus caesius L. | 53 | 63 | 18 | 36 |
| Prunus padus L. | 47 | 59 | 18 | 43 |
| Sorbus aucuparia L. | 42 | 6 | 27 | 21 |
| Corylus avelana L. | 32 | 59 | 0 | 21 |
| Euonymus europaeus L. | 21 | 12 | 9 | 64 |
| Lonicera xylosteum L. | 16 | 41 | 9 | 14 |
| Rhamnus cathartica L. | 11 | 29 | 0 | 50 |
| Herbaceous species | ||||
| Veronica chamaedrys L. | 58 | 31 | 64 | 36 |
| Geum urbanum A. Gray | 53 | 31 | 18 | 21 |
| Galium aparine L. | 42 | 19 | 18 | 7 |
| Aegopodium podagraria L. | 32 | 44 | 9 | 36 |
| Carex hirta L. | 26 | 19 | 45 | 7 |
| Calamagrostis epigejos (L.) Roth. | 21 | 13 | 55 | 29 |
| Taraxacum officinale F.H. Wigg. | 21 | 6 | 55 | 14 |
| Vicia cracca L. | 21 | 0 | 45 | 36 |
| Equisetum arvense L. | 11 | 31 | 55 | 43 |
| Dactylis glomerata L. | 11 | 0 | 45 | 21 |
| Poa pratensis L. | 11 | 0 | 45 | 43 |
| Agrimonia eupatoria L. | 0 | 6 | 0 | 43 |
| Achillea millefolium L. | 0 | 0 | 27 | 43 |
| Briza media L. | 0 | 0 | 0 | 43 |
| Medicago falcata L. | 0 | 0 | 9 | 43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Petrulaitis, L.; Gudžinskas, Z. Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania. Diversity 2023, 15, 107. https://doi.org/10.3390/d15010107
AMA Style
Petrulaitis L, Gudžinskas Z. Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania. Diversity. 2023; 15(1):107. https://doi.org/10.3390/d15010107
Chicago/Turabian StylePetrulaitis, Lukas, and Zigmantas Gudžinskas. 2023. "Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania" Diversity 15, no. 1: 107. https://doi.org/10.3390/d15010107
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.