
A Possible Perspective of Recultivation with Arbuscular Mycorrhiza-Inoculated Drought-Tolerant Herbaceous Plants

 ,
,
Abstract
:1. Introduction
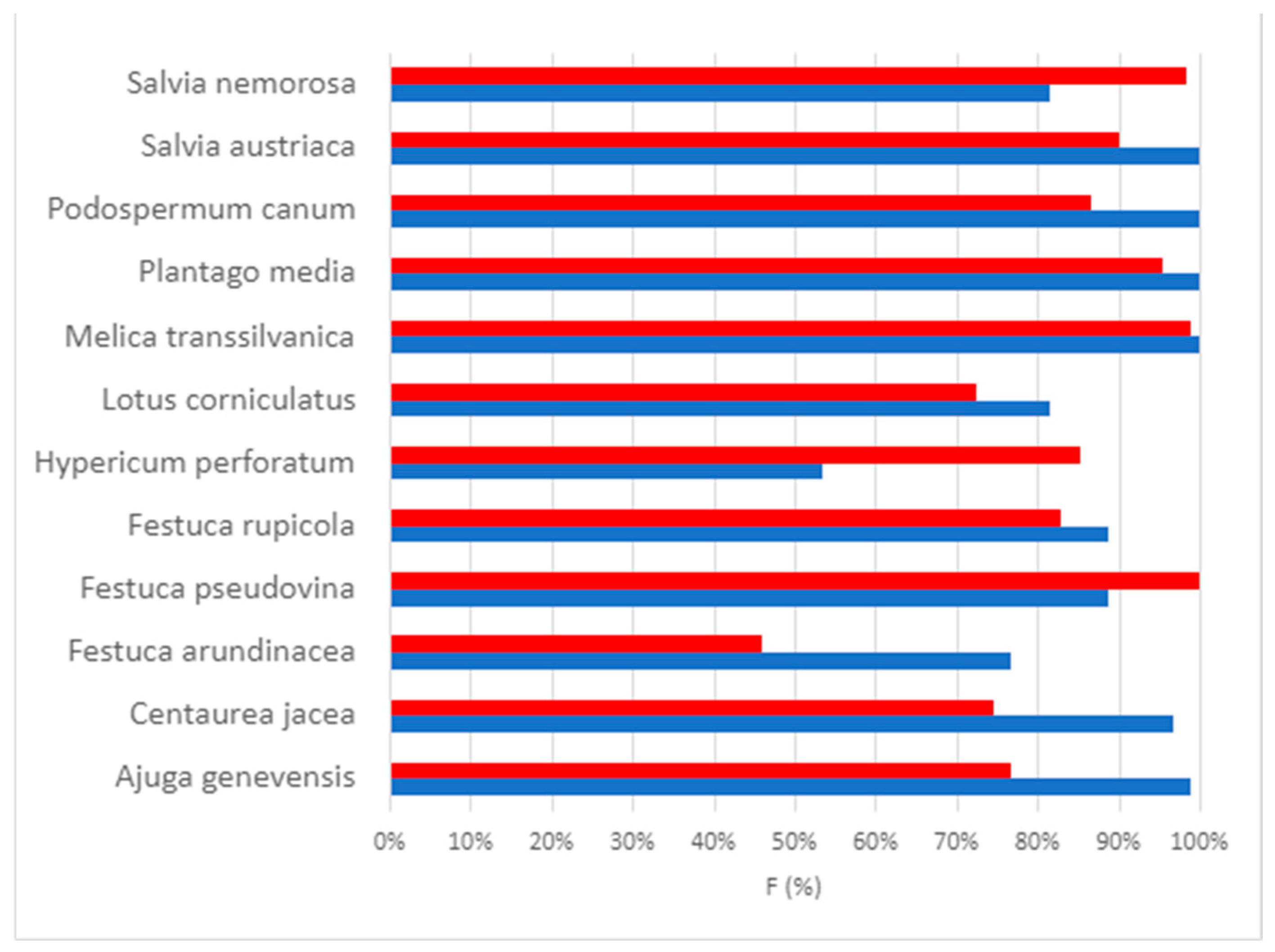
2. Results
2.1. Germination Test
2.2. Plant Morphometric Analysis
2.3. Root Investigations
3. Discussion
4. Materials and Methods
4.1. Experimental Setups and Germination Experiment
4.2. Morphological Studies
4.3. Investigation of Mycorrhizal Colonization
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Name | F% | M% | a% | A% | v% | V% | |
|---|---|---|---|---|---|---|---|
| Ajuga genevensis (C) | Mean | 98.9 | 90.0 | 41.2 | 36.7 | 13.5 | 12.2 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 1.9 | 3.9 | 16.8 | 14.0 | 7.5 | 6.8 | |
| Std. Error of Mean | 1.1 | 2.3 | 9.7 | 8.1 | 4.3 | 3.9 | |
| Ajuga genevensis (M) | Mean | 76.7 | 51.1 | 19.8 | 18.7 | 36.3 | 8.1 |
| N | 2 | 2 | 2 | 2 | 2 | 2 | |
| Std. Deviation | 33.0 | 61.0 | 28.1 | 26.4 | 34.2 | 4.7 | |
| Std. Error of Mean | 23.3 | 43.1 | 19.8 | 18.7 | 24.2 | 3.3 | |
| Centaurea jacea (C) | Mean | 96.7 | 74.1 | 38.7 | 29.1 | 33.0 | 23.7 |
| N | 2 | 2 | 2 | 2 | 2 | 2 | |
| Std. Deviation | 4.7 | 22.5 | 3.6 | 11.4 | 6.3 | 2.7 | |
| Std. Error of Mean | 3.3 | 15.9 | 2.6 | 8.1 | 4.5 | 1.9 | |
| Centaurea jacea (M) | Mean | 74.5 | 35.0 | 19.5 | 5.6 | 54.2 | 13.5 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 21.6 | 26.6 | 10.0 | 3.6 | 35.5 | 9.2 | |
| Std. Error of Mean | 12.5 | 15.4 | 5.8 | 2.1 | 20.5 | 5.3 | |
| Festuca arundinacea (C) | Mean | 76.7 | 47.9 | 16.4 | 10.0 | 28.2 | 12.2 |
| N | 4 | 4 | 4 | 4 | 4 | 4 | |
| Std. Deviation | 38.0 | 32.3 | 16.7 | 10.4 | 18.9 | 13.1 | |
| Std. Error of Mean | 19.0 | 16.2 | 8.4 | 5.2 | 9.4 | 6.6 | |
| Festuca arundinacea (M) | Mean | 46.1 | 13.5 | 17.4 | 2.2 | 38.3 | 8.6 |
| N | 2 | 2 | 2 | 2 | 2 | 2 | |
| Std. Deviation | 32.3 | 12.9 | 1.9 | 2.0 | 54.2 | 12.2 | |
| Std. Error of Mean | 22.8 | 9.1 | 1.3 | 1.4 | 38.3 | 8.6 | |
| Festuca pseudovina (C) | Mean | 88.9 | 48.0 | 13.8 | 5.9 | 54.1 | 25.7 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 10.2 | 19.6 | 6.9 | 2.7 | 3.3 | 9.3 | |
| Std. Error of Mean | 5.9 | 11.3 | 4.0 | 1.6 | 1.9 | 5.4 | |
| Festuca pseudovina (M) | Mean | 100.0 | 77.4 | 4.6 | 3.7 | 4.0 | 3.2 |
| N | 2 | 2 | 2 | 2 | 2 | 2 | |
| Std. Deviation | 0.0 | 3.2 | 5.7 | 4.5 | 5.6 | 4.5 | |
| Std. Error of Mean | 0.0 | 2.3 | 4.0 | 3.2 | 4.0 | 3.2 | |
| Festuca rupicola (C) | Mean | 88.9 | 60.3 | 19.1 | 11.7 | 17.1 | 6.6 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 19.2 | 40.3 | 19.1 | 18.0 | 16.9 | 7.7 | |
| Std. Error of Mean | 11.1 | 23.3 | 11.0 | 10.4 | 9.7 | 4.4 | |
| Festuca rupicola (M) | Mean | 83.0 | 58.6 | 22.1 | 18.7 | 33.2 | 14.0 |
| N | 4 | 4 | 4 | 4 | 4 | 4 | |
| Std. Deviation | 19.9 | 36.9 | 32.0 | 31.7 | 22.0 | 3.1 | |
| Std. Error of Mean | 9.9 | 18.4 | 16.0 | 15.9 | 11.0 | 1.5 | |
| Hypericum perforatum (C) | Mean | 53.4 | 19.7 | 6.7 | 2.6 | 60.4 | 5.6 |
| N | 2 | 2 | 2 | 2 | 2 | 2 | |
| Std. Deviation | 36.6 | 26.7 | 9.4 | 3.6 | 47.2 | 6.8 | |
| Std. Error of Mean | 25.9 | 18.9 | 6.7 | 2.6 | 33.4 | 4.8 | |
| Hypericum perforatum (M) | Mean | 85.3 | 60.2 | 29.2 | 18.4 | 41.6 | 21.6 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 12.8 | 19.8 | 15.5 | 13.2 | 26.4 | 10.3 | |
| Std. Error of Mean | 7.4 | 11.4 | 8.9 | 7.6 | 15.2 | 5.9 | |
| Lotus corniculatus (C) | Mean | 81.7 | 57.0 | 55.8 | 43.7 | 33.6 | 8.2 |
| N | 2 | 2 | 2 | 2 | 2 | 2 | |
| Std. Deviation | 25.9 | 46.0 | 51.7 | 55.1 | 47.5 | 11.6 | |
| Std. Error of Mean | 18.3 | 32.5 | 36.5 | 39.0 | 33.6 | 8.2 | |
| Lotus corniculatus (M) | Mean | 72.5 | 35.9 | 16.3 | 9.4 | 45.1 | 8.0 |
| N | 4 | 4 | 4 | 4 | 4 | 4 | |
| Std. Deviation | 21.3 | 32.2 | 16.4 | 15.4 | 38.4 | 6.0 | |
| Std. Error of Mean | 10.7 | 16.1 | 8.2 | 7.7 | 19.2 | 3.0 | |
| Melica transsilvanica (C) | Mean | 100.0 | 88.2 | 33.8 | 29.5 | 26.0 | 22.8 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 0.0 | 3.7 | 19.9 | 17.0 | 12.8 | 11.2 | |
| Std. Error of Mean | 0.0 | 2.1 | 11.5 | 9.8 | 7.4 | 6.4 | |
| Melica transsilvanica (M) | Mean | 98.9 | 82.9 | 5.3 | 4.6 | 6.1 | 5.3 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 1.9 | 9.5 | 8.5 | 7.5 | 5.5 | 5.1 | |
| Std. Error of Mean | 1.1 | 5.5 | 4.9 | 4.3 | 3.2 | 2.9 | |
| Plantago media (C) | Mean | 100.0 | 88.4 | 37.7 | 33.3 | 8.3 | 7.4 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 0.0 | 3.9 | 20.2 | 18.1 | 1.2 | 1.2 | |
| Std. Error of Mean | 0.0 | 2.2 | 11.7 | 10.5 | 0.7 | 0.7 | |
| Plantago media (M) | Mean | 95.6 | 62.3 | 47.0 | 34.3 | 29.8 | 14.4 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 7.7 | 31.8 | 24.3 | 27.2 | 21.2 | 5.5 | |
| Std. Error of Mean | 4.4 | 18.4 | 14.1 | 15.7 | 12.2 | 3.2 | |
| Podospermum canum (C) | Mean | 100.0 | 76.6 | 59.4 | 45.1 | 37.6 | 28.1 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 0.0 | 31.2 | 21.0 | 26.5 | 6.4 | 11.0 | |
| Std. Error of Mean | 0.0 | 18.0 | 12.1 | 15.3 | 3.7 | 6.4 | |
| Podospermum canum (M) | Mean | 86.7 | 77.1 | 52.0 | 37.3 | 4.1 | 3.7 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 23.1 | 26.7 | 22.5 | 15.1 | 7.1 | 6.5 | |
| Std. Error of Mean | 13.3 | 15.4 | 13.0 | 8.7 | 4.1 | 3.7 | |
| Salvia austriaca (C) | Mean | 100.0 | 89.4 | 48.6 | 43.5 | 7.4 | 6.5 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 0.0 | 9.6 | 16.0 | 16.1 | 3.7 | 3.5 | |
| Std. Error of Mean | 0.0 | 5.6 | 9.2 | 9.3 | 2.1 | 2.0 | |
| Salvia austriaca (M) | Mean | 90.0 | 72.9 | 9.5 | 8.6 | 23.4 | 12.6 |
| N | 3 | 3 | 3 | 3 | 3 | 3 | |
| Std. Deviation | 17.3 | 30.3 | 13.2 | 11.9 | 22.7 | 6.2 | |
| Std. Error of Mean | 10.0 | 17.5 | 7.6 | 6.9 | 13.1 | 3.6 | |
| Salvia nemorosa (C) | Mean | 81.7 | 38.1 | 5.7 | 2.6 | 29.9 | 6.0 |
| N | 2 | 2 | 2 | 2 | 2 | 2 | |
| Std. Deviation | 11.8 | 25.4 | 3.3 | 2.7 | 42.3 | 8.5 | |
| Std. Error of Mean | 8.3 | 18.0 | 2.3 | 1.9 | 29.9 | 6.0 | |
| Salvia nemorosa (M) | Mean | 98.3 | 63.1 | 33.5 | 25.6 | 34.5 | 20.0 |
| N | 4 | 4 | 4 | 4 | 4 | 4 | |
| Std. Deviation | 3.3 | 26.9 | 26.6 | 23.5 | 10.4 | 5.1 | |
| Std. Error of Mean | 1.7 | 13.4 | 13.3 | 11.7 | 5.2 | 2.6 |
References
- Mallik, A.U.; Karim, M.N. Roadside revegetation with native plants: Experimental seeding and transplanting of stem cuttings. Appl. Veg. Sci. 2008, 11, 547–554. [Google Scholar] [CrossRef]
- Turnau, K.; Orlowska, E.; Ryszka, P.; Zubek, S.; Anielska, T.; Gawronski, S.; Jurkie-wicz, A. Role of mycorrhizal fungi in phytoremediation and toxicity monitor-ing of heavy metal rich industrial wastes in Southern Poland. In Soil and Water Pollution Monitoring, Protection and Remediation; Springer: Dordrecht, The Netherlands, 2006; pp. 533–551. [Google Scholar]
- Berger, J.J. Ecological restoration and nonindigenous plant species: A review. Restor. Ecol. 1993, 1, 74–82. [Google Scholar] [CrossRef]
- Miransari, M. Contribution of arbuscular mycorrhizal symbiosis to plant growth under different types of soil stress. Plant Biol. 2010, 12, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Borhidi, A. Magyarország Növénytársulásai; Akadémiai Kiadó: Budapest, Hungary, 2003; p. 610. [Google Scholar]
- Partzsch, M.; Faulhaber, M.; Meier, T. The effect of the dominant grass Festuca rupicola on the establishment of rare forbs in semi-dry grasslands. Folia Geobot. 2018, 53, 103–113. [Google Scholar] [CrossRef]
- Valkó, O.; Tóth, K.; Kelemen, A.; Miglécz, T.; Radócz, S.; Sonkoly, J.; Tóthmérész, B.; Török, P.; Deák, B. Cultural heritage and biodiversity conservation–plant introduction and practical restoration on ancient burial mounds. Nat. Conserv. 2018, 24, 65–80. [Google Scholar] [CrossRef]
- Szirmai, O.; Saláta, D.; Benedek, L.K.; Czóbel, S. Investigation of the secondary succession of abandoned areas from different cultivation in the Pannonian biogeographic region. Agronomy 2022, 12, 773. [Google Scholar] [CrossRef]
- Kis, S. Adatok a vasútmenti pionír élőhelyek flórájához a Tiszántúlon. Kitaibelia 2022, 27, 86–101. [Google Scholar] [CrossRef]
- Westbury, D.B.; Woodcock, B.A.; Harris, S.J.; Brown, V.K.; Potts, S.G. The effects of seed mix and management on the abundance of desirable and pernicious unsown species in arable buffer strip communities. Weed Res. 2008, 48, 113–123. [Google Scholar] [CrossRef]
- Landis, T.D.; Wilkinson, K.M.; Steinfeld, D.; Riley, S.A.; Fekaris, G. Roadside revegetation of forest highways: New applications for native plants. Nativ. Plants J. 2005, 6, 297–305. [Google Scholar] [CrossRef]
- Török, P.; Brudvig, L.A.; Kollmann, J.; NPrice, J.; Tóthmérész, B. The present and future of grassland restoration. Restor. Ecol. 2021, 29, e13378. [Google Scholar] [CrossRef]
- Török, P.; Vida, E.; Deák, B.; Lengyel, S.; Tóthmérész, B. Grassland restoration on former croplands in Europe: An assessment of applicability of techniques and costs. Biodivers. Conserv. 2011, 20, 2311–2332. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010; p. 605. [Google Scholar]
- Harley, J.L.; Harley, E.L. A check-list of mycorrhiza in the British flora. New Phytol. 1987, 105, 1–102. [Google Scholar] [CrossRef]
- Kovács, M.G. Magyarországi növények mikorrhizáltsági vizsgálatainak összefoglalása. Kitaibelia 2008, 13, 62–73. [Google Scholar]
- Harrison, M.J. The arbuscular mycorrhizal symbiosis. In Plant-Microbe Interactions; Springer: Boston, FL, USA, 1997; pp. 1–34. [Google Scholar]
- Salomon, M.J.; Demarmels, R.; Watts-Williams, S.J.; McLaughlin, M.J.; Kafle, A.; Ketelsen, C.; Soupir, A.; Bücking, H.; Cavagnaro, T.R.; van der Heijden, M.G.A. Global evaluation of commercial arbuscular mycorrhizal inoculants under greenhouse and field conditions. Appl. Soil Ecol. 2022, 169, 104225. [Google Scholar] [CrossRef]
- Turnau, K.; Gawroński, S.; Ryszka, P.; Zook, D. Mycorrhizal-based phytostabilization of Zn–Pb tailings: Lessons from the Trzebionka mining works (Southern Poland). In Bio-Geo Interactions in Metal-Contaminated Soils; Springer: Berlin/Heidelberg, Germany, 2012; pp. 327–348. [Google Scholar]
- Eschen, R.; Müller-Schärer, H.; Schaffner, U. Aboveground environment type, soil nutrient content and arbuscular mycorrhizal fungi explain establishment success of Centaurea jacea on ex-arable land and in late-successional grasslands. Plant Soil 2009, 322, 115–123. [Google Scholar] [CrossRef]
- Zoltán, L.; Kalapos, T.; Endresz, G. Négy adventív lágyszárú növényfaj mikorrhizáltsága bolygatott élőhelyeken. Bot. Közlemények 2014, 101, 39–49. [Google Scholar]
- Seifert, E.K.; Bever, J.D.; Maron, J.L. Evidence for the evolution of reduced mycorrhizal dependence during plant invasion. Ecology 2009, 90, 1055–1062. [Google Scholar] [CrossRef]
- Bennett, A.E.; Meek, H.C. The influence of arbuscular mycorrhizal fungi on plant reproduction. J. Chem. Ecol. 2020, 46, 707–721. [Google Scholar] [CrossRef]
- Richter, J.; Baltruschat, H.; Kabrodt, K.; Schellenberg, I. Impact of arbuscular mycorrhiza on the St. John’s wort (Hypericum perforatum) wilt disease induced by Colletotrichum cf. gloeosporioides. J. Plant Dis. Prot. 2011, 118, 109–118. [Google Scholar] [CrossRef]
- Zarei, M.; Saleh-Rastin, N.; Jouzani, G.S.; Savaghebi, G.; Buscot, F. Arbuscular mycorrhizal abundance in contaminated soils around a zinc and lead deposit. Eur. J. Soil Biol. 2008, 44, 381–391. [Google Scholar] [CrossRef]
- Shao, Y.; Jiang, S.; Peng, H.; Li, H.; Li, P.; Jiang, R.; Fang, W.; Chen, T.; Jiang, G.; Yang, T.; et al. Indigenous and commercial isolates of arbuscular mycorrhizal fungi display differential effects in Pyrus betulaefolia roots and elicit divergent transcriptomic and metabolomic responses. Front. Plant Sci. 2023, 13, 1040134. [Google Scholar] [CrossRef] [PubMed]
- Füzy, A.; Biró, B.; Tóth, T.; Hildebrandt, U.; Bothe, H. Drought, but not salinity, determines the apparent effectiveness of halophytes colonized by arbuscular mycorrhizal fungi. J. Plant Physiol. 2008, 165, 1181–1192. [Google Scholar] [CrossRef] [PubMed]
- Boldt-Burisch, K.; Naeth, M.A. Mycorrhization affects root distribution of Lotus corniculatus and Calamagrostis epigeios in a nutrient poor heterogeneous soil in a rhizotron experiment. Rhizosphere 2017, 4, 36–47. [Google Scholar] [CrossRef]
- Engelmoer, D.J.; Behm, J.E.; Toby Kiers, E. Intense competition between arbuscular mycorrhizal mutualists in an in vitro root microbiome negatively affects total fungal abundance. Mol. Ecol. 2014, 23, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Thonar, C.; Frossard, E.; Šmilauer, P.; Jansa, J. Competition and facilitation in synthetic communities of arbuscular mycorrhizal fungi. Mol. Ecol. 2014, 23, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Király, G. (Ed.) Új Magyar Füvészkönyv; Magyarország hajtásos növényei; Határozókulcsok; Aggteleki Nemzeti Park Igazgatóság: Jósvafő, Hungary, 2009; p. 616. [Google Scholar]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.R.S.; Piché, Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [PubMed]
- Vierheilig, H.; Schweiger, P.; Brundrett, M. An overview of methods for the detection and observation of arbuscular mycorrhizal fungi in roots. Physiol. Plant. 2005, 125, 393–404. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Estimation of V. A. mycorrhizal infection levels: Research for methods having a functional significance. In Proceedings of the 1st European Symposium, Physiological and Genetical Aspects of Mycorrhizae, Dijon, France, 1–5 July 1985; Station d’Amelioration des Plantes: Paris, France, 1986; pp. 217–221. [Google Scholar]







| Taxon Name | Treatment | Origin | Aver. Germ. per 20 Seeds | SD Germ. per 20 Seeds | Germ. Perc. (%) | Pairwise p-Value < 0.05 | |
|---|---|---|---|---|---|---|---|
| 1. | Ajuga genevensis L. | C | Abádszalók 47.401823, 20.677957 | 1.0 | 1.00 | 5 | 1 |
| M | 1.0 | 1.73 | 5 | ||||
| 2. | Centaurea jacea L. | C | Abádszalók | 12.0 | 3.00 | 60 | 0.208 |
| M | 47.416913, 20.693190 | 9.0 | 1.73 | 45 | |||
| 3. | Echium vulgare L. | C | Abádszalók | 3.3 | 2.08 | 17 | 0.115 |
| M | 47.412539, 20.689428 | 6.3 | 1.53 | 32 | |||
| 4. | Festuca arundinacea Schreb. | C | Törökszentmiklós | 8.7 | 1.15 | 43 | 0.039 |
| M | 47.123444, 20.480192 | 6.0 | 1.00 | 30 | |||
| 5. | Festuca pseudovina Hack. | C | Törökszentmiklós | 18.0 | 2.65 | 90 | 0.192 |
| M | 47.124260, 20.482027 | 15.0 | 2.00 | 75 | |||
| 6. | Festuca rupicola Heuff. | C | Törökszentmiklós | 15.3 | 3.51 | 77 | 0.180 |
| M | 47.124995, 20.483616 | 11.0 | 3.00 | 55 | |||
| 7. | Filipendula vulgaris Moench | C | Abádszalók | 8.0 | 3.00 | 40 | 1 |
| M | 47.404759, 20.679292 | 8.0 | 5.00 | 40 | |||
| 8. | Hypericum perforatum L. | C | Abádszalók | 1.0 | 1.00 | 5 | 0.016 |
| M | 47.417316, 20.692507 | 3.7 | 0.58 | 18 | |||
| 9. | Lotus corniculatus L. | C | Kunmadaras | 4.0 | 2.00 | 20 | 0.609 |
| M | 47.430432, 20.860312 | 3.3 | 0.58 | 17 | |||
| 10. | Melica transsilvanica Schur | C | Törökszentmiklós | 11.7 | 4.04 | 58 | 1 |
| M | 47.124800, 20.482981 | 11.7 | 5.51 | 58 | |||
| 11. | Plantago media L. | C | Törökszentmiklós | 6.3 | 1.15 | 32 | 0.055 |
| M | 47.122457, 20.478377 | 2.3 | 2.31 | 18 | |||
| 12. | Podospermum canum (C.A. Mey.) Griseb. | C | Kunmadaras | 10.0 | 1.00 | 50 | 0.034 |
| M | 47.430441, 20.860406 | 6.7 | 1.53 | 33 | |||
| 13. | Pseudolysimachion spicatum (L.) Opiz | C | Abádszalók | 1.0 | 0.00 | 5 | 0.374 |
| M | 47.417119, 20.692839 | 1.3 | 0.58 | 7 | |||
| 14. | Salvia austriaca Jacq. | C | Tiszafüred | 7.3 | 0.58 | 37 | 0.013 |
| M | 47.591402, 20.851583 | 9.3 | 0.58 | 47 | |||
| 15. | Salvia nemorosa L. | C | Törökszentmiklós | 9.7 | 2.52 | 48 | 0.011 |
| M | 47.123603, 20.480440 | 2.7 | 1.15 | 13 | |||
| 16. | Salvia pratensis L. | C | Tiszaszentimre | 6.3 | 0.58 | 32 | 1 |
| M | 47.468523, 20.804884 | 6.3 | 1.53 | 32 | |||
| 17. | Silene viscosa (L.) Pers | C | Abádszalók | 9.0 | 5.29 | 45 | 0.272 |
| M | 47.416194, 20.690254 | 4.3 | 3.51 | 22 | |||
| 18. | Thymus glabrescens Willd. | C | Abádszalók | 0.7 | 0.58 | 3 | 0.519 |
| M | 47.416850, 20.690566 | 0.3 | 0.58 | 2 | |||
| 19. | Trifolium pratense L. | C | Törökszentmiklós | 1.0 | 1.00 | 5 | 0.561 |
| M | 47.123371, 20.480228 | 1.7 | 1.53 | 8 | |||
| 20. | Verbascum phoeniceum L. | C | Abádszalók | 0.7 | 0.58 | 3 | 0.116 |
| M | 47.416492, 20.690413 | 0.0 | 0.00 | 0 | |||
| Taxon Name | Treatment | No. of Specimen | Average Stem Length (cm) | SD Stem Length | p-Value < 0.05 | ||
|---|---|---|---|---|---|---|---|
| 1. | Festuca arundinacea | C | 26 | 61.0 | 13.4 | 0.535 | |
| M | 18 | 51.5 | 11.7 | ||||
| 2. | Festuca rupicola | C | 46 | 30.6 | 4.9 | 0.462 | |
| M | 33 | 28.5 | 7.2 | ||||
| 3. | Festuca pseudovina | C | 54 | 20.5 | 4.1 | 0.286 | |
| M | 45 | 26.8 | 5.1 | ||||
| 4. | Melica transsilvanica | C | 35 | 25.4 | 5.5 | 0.668 | |
| M | 35 | 26.7 | 5.0 | ||||
| 5. | Lotus corniculatus | C | 12 | 29.7 | 12.5 | 0.630 | |
| M | 10 | 33.9 | 9.2 | ||||
| 6. | Hypericum perforatum | C | 3 | 14.2 | 10.9 | 0.700 | |
| M | 1 | 19.1 | 10.2 | ||||
| Taxon Name | Treatment | No. of Specimen | Average Leaf No. | p-Value < 0.05 | SD Leaf No | Average Leaf Length (cm) | p-Value < 0.05 | SD Leaf Length | Average Leaf Width (cm) | p-Value < 0.05 | Std. Deviation of Leaf Width | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. | Ajuga genevensis | C | 3 | 6 | 0.116 | 0.6 | 4.3 | 0.862 | 0.7 | 1.5 | 0.775 | 1.3 |
| M | 3 | 7 | 0 | 4.6 | 0.8 | 1.5 | 0.6 | |||||
| 2. | Centaurea jacea | C | 36 | 7 | 0.236 | 1.7 | 15.7 | 6.1 | 1.8 | 0.7 | ||
| M | 27 | 7 | 1.7 | 15.5 | 0.823 | 5.6 | 1.9 | 0.811 | 0.6 | |||
| 3. | Echium vulgare | C | 10 | 8 | 0.000 | 5.5 | 8.5 | 0.011 | 4 | 1.3 | 0.647 | 0.7 |
| M | 19 | 4 | 1.3 | 3.7 | 2.6 | 0.8 | 0.3 | |||||
| 4. | Filipendula vulgaris | C | 25 | 9 | 0.719 | 6.2 | 3.7 | 0.142 | 2.1 | 1.4 | 1 | 0.6 |
| M | 26 | 7 | 2.2 | 4.9 | 2.3 | 1.7 | 0.5 | |||||
| 5. | Plantago media | C | 18 | 6 | 0.000 | 2.3 | 8.1 | 0.021 | 4.8 | 1.6 | 0.385 | 0.6 |
| M | 10 | 11 | 4.9 | 13.7 | 5.6 | 2.2 | 2.2 | |||||
| 6. | Podospermum canum | C | 30 | 9 | 0.000 | 4.3 | 16.2 | 0.032 | 4.9 | 0.4 | 0.164 | 0.2 |
| M | 20 | 5 | 1.9 | 10.8 | 3.2 | 0.6 | 0.6 | |||||
| 7. | Pseudolysimachion spicatum | C | 3 | 16 | 0.607 | 6.1 | 5.8 | 0.189 | 1.7 | 2.4 | 0.487 | 0.7 |
| M | 4 | 14 | 3.7 | 6 | 0.7 | 2.3 | 0.6 | |||||
| 8. | Salvia austriaca | C | 22 | 7 | 0.820 | 2.2 | 7.2 | 0.007 | 3.3 | 1.9 | 0.817 | 0.8 |
| M | 28 | 7 | 2 | 4.9 | 1.9 | 1.4 | 0.7 | |||||
| 9. | Salvia nemorosa | C | 29 | 19 | 0.061 | 8.7 | 5.5 | 0.431 | 2.6 | 1.9 | 0.390 | 0.9 |
| M | 8 | 14 | 7 | 5.7 | 3.2 | 1.6 | 0.9 | |||||
| 10. | Salvia pratensis | C | 17 | 7 | 0.001 | 14.7 | 5.6 | 0.515 | 2.6 | 1.8 | 0.797 | 0.9 |
| M | 19 | 9 | 2 | 9 | 4.6 | 2.3 | 1.2 | |||||
| 11. | Silene viscosa | C | 27 | 10 | 0.206 | 3.7 | 6.3 | 0.422 | 3 | 1.8 | 0.580 | 0.8 |
| M | 13 | 11 | 4.3 | 6.3 | 2.8 | 1.1 | 0.9 |
| Elements | Soil Depth 0–20 cm | Soil Depth 20–40 cm |
|---|---|---|
| pH (KCl) | 6.22 pH unit | 6.42 pH unit |
| humus content | 4.15 m/m% | 2.51 m/m% |
| all water-soluble salts | 0.05 m/m% | 0.05 m/m% |
| Na | 38.7 mg/kg | 25.2 mg/kg |
| Mg | 88.3 mg/kg | 69.1 mg/kg |
| S | 5.51 mg/kg | 4.95 mg/kg |
| Zn | 2.48 mg/kg | 1.08 mg/kg |
| Mn | 136 mg/kg | 81.3 mg/kg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papp, L.; Habtemariam, A.A.; Brandt, S.; Cseh, P.; Heller, Á.; Péter, B.; Szakály, Á.P.; Kiszel, P.; Codogno, B.; Bratek, Z.; et al. A Possible Perspective of Recultivation with Arbuscular Mycorrhiza-Inoculated Drought-Tolerant Herbaceous Plants. Plants 2023, 12, 4088. https://doi.org/10.3390/plants12244088
Papp L, Habtemariam AA, Brandt S, Cseh P, Heller Á, Péter B, Szakály ÁP, Kiszel P, Codogno B, Bratek Z, et al. A Possible Perspective of Recultivation with Arbuscular Mycorrhiza-Inoculated Drought-Tolerant Herbaceous Plants. Plants. 2023; 12(24):4088. https://doi.org/10.3390/plants12244088
Chicago/Turabian StylePapp, László, Akale Assamere Habtemariam, Sára Brandt, Péter Cseh, Ádám Heller, Balázs Péter, Ágnes Pappné Szakály, Péter Kiszel, Borbála Codogno, Zoltán Bratek, and et al. 2023. "A Possible Perspective of Recultivation with Arbuscular Mycorrhiza-Inoculated Drought-Tolerant Herbaceous Plants" Plants 12, no. 24: 4088. https://doi.org/10.3390/plants12244088