
Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia

Silva Tarouca Research Institute for Landscape and Ornamental Gardening, Public Research Institute, Květnové náměstí 391, 252 43 Průhonice, Czech Republic
*
Author to whom correspondence should be addressed.
Plants 2023, 12(3), 488; https://doi.org/10.3390/plants12030488
Submission received: 7 November 2022
/
Revised: 13 January 2023
/
Accepted: 14 January 2023
/
Published: 20 January 2023
(This article belongs to the Special Issue Application of Biotechnology to Woody Propagation)
Abstract
:Members of the genus Sorbus are the only endemic tree species that occur in Czechia. They are important components of endangered plant communities. Their natural regeneration is usually problematic because of their mode of reproduction and because they can survive in rare populations with small numbers of individuals. The aim of this study was to develop a successful micropropagation protocol for selected Sorbus species, of which two are endemic (S. gemella and S. omissa) and two are hybrid (S. × abscondita and S. × kitaibeliana). We found significant differences in shoot induction and rooting ability between the Sorbus species under study. With the exception of S. × abscondita, N6-benzyladenine had a significantly greater effect on shoot regeneration, both in terms of shoot number and total shoot length, than meta-topolin. Root induction was key to the successful micropropagation of the Sorbus species studied. Our results show that four Sorbus species can be successfully rooted under ex vitro conditions, without a rooting powder treatment in a steamed peat-perlite substrate. Auxin-untreated microcuttings of S. gemella, S. × kitaibeliana and S. omissa, but not S. × abscondita, rooted better than ones treated with indole-3-butyric acid. This is the first time a micropropagation protocol for S. omissa, S. × abscondita and S. × kitaibeliana has been published.
1. Introduction
In the flora of Europe, the genus Sorbus L. is one of the most taxonomically challenging groups of woody plants. Taxonomical complications stem chiefly from the vast morphological variability and huge genetic diversity of the genus, which is generated by frequent interspecific hybridization, genome duplication and subsequent stabilization of offspring by apomixis [1,2,3,4,5]. Furthermore, diverse opinions on taxonomic treatment [6,7] and a tangled nomenclature still cause disagreements and misinterpretations.
Most Sorbus species are polyploids that are locally distributed in geographically clearly defined areas, and some are stenoendemics occurring in a single locality with one population or a few subpopulations. All polyploid apomictic Sorbus taxa in Central Europe are derived from hybrids of members of Sorbus subgen. Aria Pers. (including diploid S. aria (L.) Crantz and S. umbellata (Desf.) Fritsch and polyploid apomictic taxa) or from hybrids between S. subgen. Aria and members of one of the three following subgenera: S. subgen. Sorbus (diploid S. aucuparia L.), S. subgen. Torminaria (DC.) Reichenb. (diploid S. torminalis (L.) Crantz) and S. subgen. Chamaemespilus (Medik.) K. Koch (diploid S. chamaemespilus (L.) Crantz). Furthermore, the same parental combination can polytopically form various apomictic, morphologically homogeneous and genetically unique lines which are classified in modern taxonomy as separate species (i.e., microspecies or agamospecies). As well as the species mentioned above, another diploid, S. domestica L. (S. subgen. Cormus (Spach) Duch.), occurs in Europe. Diploids are sexual and are rather widely distributed. In rare instances, they form diploid, sexual and variable offspring (S. aria × S. aucuparia = S. × thuringiaca (Nyman) Fritsch, S. aria × S. torminalis = S. × decipiens (Bechst.) Petz. & Kirchn., S. × ambigua (Decne.) Beck = S. aria × S. chamaemespilus).
According to current knowledge, there are 189 Sorbus species in Europe [8], but taxonomical research using contemporary biosystematic methods is still revealing novelties in this field [5,9,10,11,12], and the number of species keeps increasing. Most of them are threatened endemics listed in the European Red List of Trees [13]. In Czechia, there are 21 native Sorbus species, of which 13 are endemics and three are subendemics (S. cucullifera M. Lepší & P. Lepší; S. sudetica (Tausch) Bluff, Nees & Schauer; S. thayensis M. Lepší & P. Lepší) ([14,15]; Figure 5). Populations of (sub)endemics are not very numerous. They usually consist of only a few dozen (in extreme cases, less than 20) or, at most, hundreds of individuals. Most of them are located on sites representing residual fragments of natural biotopes in the cultural landscape, the protection of which is insufficient or even lacking altogether. In the last version of the Red List of vascular plants of the Czech Republic, they are included in the ‘threatened’ category [16].
The natural regeneration of Sorbus species limits their survival and further prosperity in natural habitats. In nature, they reproduce mainly generatively. To a limited extent, they can also spread clonally via rooted branches on avalanche paths or scree slopes (e.g., S. sudetica, and S. × decipiens; [17,18,19]) or via root offsets (S. × decipiens; own observation). They are entomogamous and mostly allogamous, so the successful development of their seeds depends on the presence of a greater number of plants within the flight range of insects. Autogamy (i.e., self-pollination) occurs only rarely (recorded in S. aucuparia and S. torminalis; [2,20,21]). As already stated, most of the species are polyploids with an apomictic mode of reproduction. All polyploid species are capable of pseudogamic reproduction [22,23,24,25]. It is therefore necessary to ensure the presence of other species (pollen donors) in order for viable seeds to form under natural conditions.
All Czech species of Sorbus are light-demanding and prefer open habitats such as rocks and screes, rock steppes, scrubs, woodland-steppes and thermophilous open pine, oak or hornbeam and ravine woodlands, as well as their fringes. As well as being semi-natural to relic vegetation, they occasionally grow in Larix decidua, Picea abies, Pinus nigra, P. sylvestris and, rarely, Robinia pseudoacacia plantations or in their clearings. The closed canopy that now prevails in woodlands in Czechia is unfavorable for the long-term survival and regular reproduction of such species. They are also potentially threatened by the disappearance of habitats, as open woodlands develop a dense canopy or are replaced by forest plantations [14].
The conventional method using seeds is most commonly used to reproduce Sorbus species [26]. Vegetative propagation by grafting or budding is used less frequently. Furthermore, this method is not suitable for endangered Sorbus species. The use of cuttings is inefficient compared to micropropagation. Plant tissue cultivation can provide a rapid method for the multiplication of endangered or threatened species when only a few stock plants are available or if the collection of plants and seeds from wild plants needs to be minimized [27].
A micropropagation protocol was first developed for economically important Sorbus species such as S. torminalis [28,29,30], S. aucuparia [29,31,32,33] and S. domestica [28,34,35,36,37,38,39,40]. These species are mainly used for wood production, as well as in the food industry and in ornamental horticulture. Research carried out in the last decade has shown that micropropagation is an important method for the propagation of rare and endemic Sorbus species or their infrageneric hybrid such as ×Malosorbus florentina [41], and then S. alnifrons, S. bohemica, S. gemella Kovanda, S. hardeggensis, S. quernea, S. rhodanthera and S. sudetica [42]. These species are important primarily from an ecological point of view, as they increase the biodiversity of woodland ecosystems. Some endemic species of Sorbus are found and grow well even in extreme conditions (with some degree of drought resistance), where common tree species do not grow. Some Sorbus species have been discovered only recently or little is known about them. In this study, we focused on the establishment of a micropropagation method of S. gemella, S. omissa Velebil, S. × abscondita Kovanda and S. × kitaibeliana Baksay & Kárpáti. The characteristics of Sorbus species used in this study and their localization are presented in Section 4, i.e., Materials and Methods.
2. Results
2.1. In Vitro Shoot Proliferation
The occurrence of explant contamination was low, ranging from 0 to 15%. Satisfactory reactivity of the explants to the in vitro cultures was achieved and reached a total of 79%. In our experiments, initial cultures of Sorbus species were established from axillary buds originating from the lower branches of donor trees. Better regeneration of shoots and roots could be expected in the subsequent stages of in vitro culture. This effect was observed in S. aucuparia, when microshoots derived from the lower branches of mature trees exhibited better shoot proliferation and rooting response than from the top branches [43]. After 6 months of cultivation, in vitro cultures of Sorbus species were stabilized.
There were significant differences (p-value < 0.05) between the Sorbus species under study in terms of the mean number of shoots per explant (Figure 1A). Sorbus × abscondita (2.71) and S. × kitaibeliana (2.35) produced significantly more shoots than S. gemella (1.88) and S. omissa (1.86).
The application of cytokinins during multiplication significantly increased shoot regeneration compared to the control (1.03; Figure 1C). A higher shoot production (4.17) was observed on the multiplication medium with N6-benzyladenine (BA) than the one with meta-topolin (mT, 1.58). The combination of each of the cytokinins with the auxin indole-3-butyric acid (IBA) had no statistical effect on the mean number of shoots.
Another growth parameter was total shoot length, in which significant differences were found between Sorbus species (Figure 1B); the response was similar to that for the mean number of shoots per explant (Figure 1A). Sorbus × abscondita (61.48 mm) and S. × kitaibeliana (53.72 mm) produced significantly longer shoots than S. omissa (38.47 mm) and S. gemella (36.07 mm).
The cytokinins also had a positive effect on total shoot length compared to the control (20.67 mm; Figure 1D). The shoots on the multiplication medium with BA were significantly longer (77.89 mm) than that with mT (39.61 mm). The combination of each of the cytokinins with the auxin (IBA) had no statistical effect on total shoot length.
The interaction between the genotype and the PGR treatment showed a different response in shoot production in three Sorbus species (Table 1). In S. × abscondita, the type of cytokinin played a significant role, and shoot formation was significantly higher on the medium with mT (3.72) or mT + IBA (3.47) than on that with BA (2.97). In S. gemella, shoot formation was significantly higher on the medium with mT + BA (2.69) compared to the control (1.11) and the IBA treatment (1.03). In S. × kitaibeliana, shoot formation was significantly higher on the medium with BA (3.42) compared to the control (1.28) and IBA treatment (1.36).
Regarding the interaction between the genotype and the PGR treatment on total shoot length, significant differences were observed in two Sorbus species (Table 2). In S. × abscondita, shoot elongation was significantly better in the mT + IBA treatment (89.22 mm) than in the combined BA + IBA treatment (54.94 mm) and the IBA treatment (39.99 mm). In S. × kitaibeliana, the combination of mT + IBA (65.50 mm) was also effective, as was the BA treatment (71.00 mm), where the total length of the shoots was significantly higher compared to the control (31.69 mm) and the IBA treatment (36.31 mm).
2.2. Ex Vitro Rooting and Acclimatization
The effects of different factors (genotype, auxin treatment/commercial rooting powder and microcutting size) on rooting were examined. All observed factors had significant effects on the rooting percentage and the mean number of roots per microcutting at different significance levels. The effects of interactions between two factors were found to be significant for genotype × auxin treatment for all growth characteristics and genotype × microcutting size for rooting percentage (Table 3).
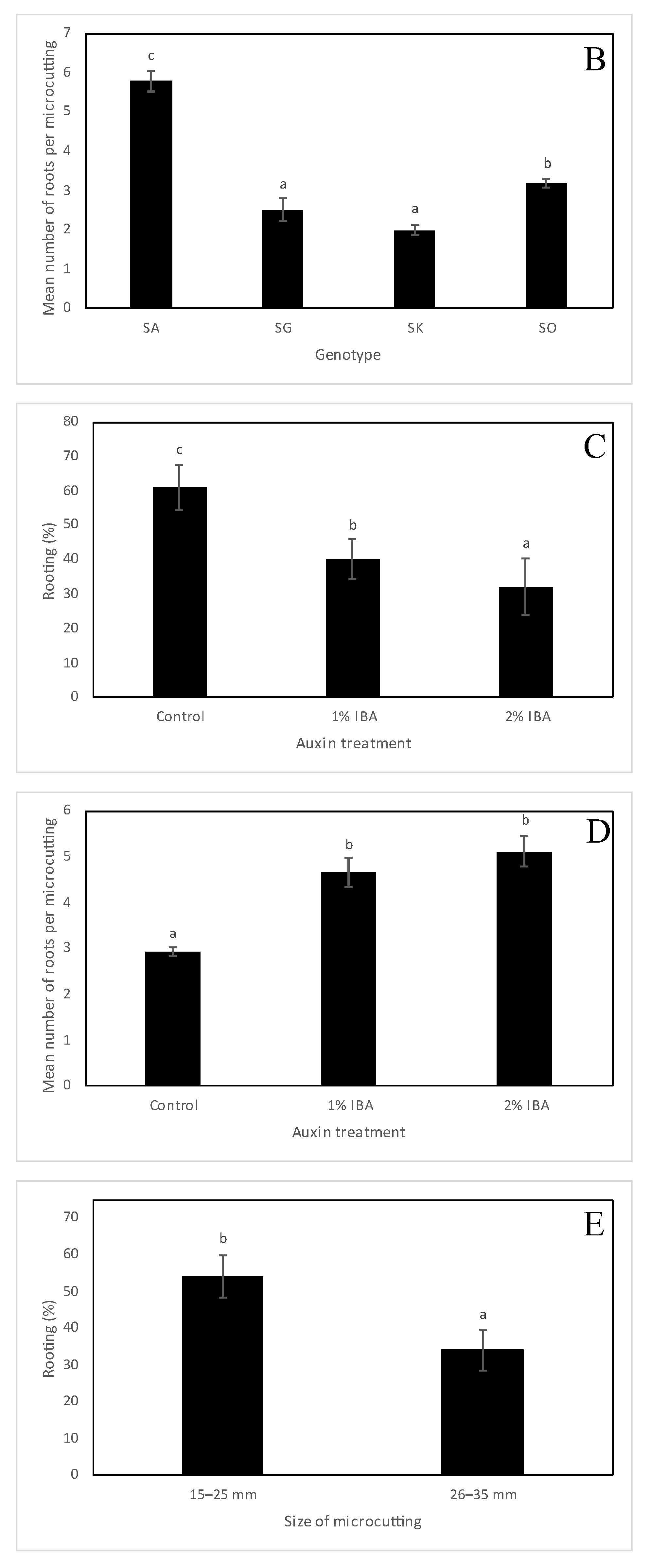
Rhizogenesis was observed in all Sorbus species. There were statistically significant differences in rooting between Sorbus × abscondita, S. gemella, S. × kitaibeliana and S. omissa (Figure 2A and Figure 3). A high rooting percentage was found in S. × abscondita (74%) and S. omissa (62%), whereas a low root capability was observed in S. × kitaibeliana (27%) and S. gemella (14%). A similar trend was also observed for the mean number of roots per microcutting (5.78, 3.18, 1.98 and 2.51, respectively; Figure 2B).
Root production also depended on the presence and the concentration of IBA in the rooting powder (Figure 2C). Auxin treatments (1% and 2% IBA) reduced the rooting percentage significantly (40% and 32%) compared to the control (61%). On the other hand, the auxin significantly increased the mean number of roots per microcutting, both in the 1% IBA treatment (4.66) and in the 2% IBA treatment (5.12.) compared with the control (2.91), see Figure 2D.
The size of the microcutting was also an important factor affecting Sorbus rooting (Figure 2E). We found that microcuttings that were 15–25 mm long rooted significantly better (54%) than microcuttings that were 26–35 mm long (34%). The opposite effect was observed for the mean number of roots per microcutting; in longer microcuttings, the formation of roots was significantly higher (4.66) than in shorter microcuttings (3.56), see Figure 2F.
The interaction between the genotype and the auxin treatment showed a different response in the rooting percentage of Sorbus × abscondita compared to other Sorbus species (Figure 2G). Only in this species was there a significant difference in rooting percentage between the control (81%) and the 2% IBA treatment (90%). The effect of auxin on the mean number of roots per microcutting was demonstrated conclusively in three Sorbus genotypes (Figure 2H). The auxin treatment increased the mean number of roots in S. × abscondita (8.0 and 6.59 with 1% and 2% IBA, respectively) and in S. omissa (1% IBA, 3.32) and had an antagonistic effect in S. gemella (2% IBA, 0.0) compared to the control. The effect of the interaction between the genotype and the size of the microcuttings on rooting percentage was significant only in S. omissa, where shorter microcuttings rooted better (84%) compared to longer microcuttings (40%; Figure 2I).
3. Discussion
In preservation programs that specialize in the propagation of endangered tree species, often only adult trees are available as a source plant. The success of micropropagation is strongly influenced by the ontogenetic and chronological age of source plants. According to [27], ontogenetic aging (physiological) refers to the phases of development that the seedling plant undergoes from embryonic to juvenile to intermediate to mature (adult), and chronological aging continues through the life of an individual plant whether as a seedling or vegetatively propagated. In vitro cultures initiated from source plants that are physiologically mature or less vital (old) or both may exhibit growth depression during micropropagation.
In vitro cultures can be successfully obtained from an adult tree in several ways: establishing cultures from plant parts showing juvenile characteristics, from mature parts after rejuvenation or from serial subcultures of explants in a hormone medium [44,45,46].
Plant growth regulators play a key role in adventitious shoot regeneration in woody species of the temperate zone [47]. The presence of a cytokinin is necessary for de novo shoot formation in woody species, and very often a cytokinin is combined with auxin at a low concentration to increase the number of shoots.
BA is an important aromatic cytokinin that is very effective at shoot induction in many plant species, including Sorbus species, and it is affordable compared to other cytokinins. On the other hand, it can cause growth abnormalities in some plant species, reduce rooting and worsen the subsequent acclimatization of plants [30,48,49]. For these reasons, new aromatic cytokinin derivatives have been the subject of intensive research [50]. One of them is mT, a natural highly active aromatic cytokinin occurring in poplar leaves [51]. Meta-topolin has been successfully used in the micropropagation of many plant species, including both herbs [49,52,53] and trees [54,55,56]. Ördögh et al. [57] published the effects of different types of cytokinin, including mT, on shoot proliferation in S. borbasii Jáv.
In our study, we compared the effectiveness of two cytokinins, BA and mT, singly or in combination, with auxin in four Sorbus species. In agreement with the results of Nikolaou et al. [37] and Jeong and Sivanesan [58], the presence of a cytokinin was essential for shoot proliferation in Sorbus species. In general, BA had a significantly higher effect on shoot regeneration, both in terms of shoot number and total shoot length, than mT (Figure 1C,D). This finding that BA increases the number of shoots more effectively than mT is consistent with the results obtained by Ördögh et al. [59] for S. redliana Kárpáti, by Malá et al. [30] for S. torminalis, by Ördögh et al. [57] for Sorbus borbasii and by Meyer et al. [60] for Hypericum L. species. On the other hand, some Sorbus species behaved differently: In S. × abscondita, shoot formation was significantly more vigorous on the multiplication medium with mT than the one with BA (Table 1 and Table 2). A similar stronger reaction to mT in shoot proliferation was observed in Ribes [61]. Likewise, Hlophe et al. [62] noticed that each Brachystelma R. Br. species differed in its response to specific cytokinins.
The combination of each of the cytokinins with the auxin IBA or the auxin alone had no significant effect on growth parameters in the Sorbus species studied (Figure 1), although a slight improvement in shoot production was observed in S. gemella and S. omissa on the medium with mT + IBA (Table 2). Similar results were obtained with S. domestica, where the addition of IBA to BA had no effect on shoot proliferation [37]. Many authors routinely used a combination of a cytokinin and the auxin IBA for proliferation without further analyses [30,41,43,57,59]. In contrast to our results, it has been reported that the combination of a cytokinin (BA) and auxin (IAA or NAA) had a synergistic effect on shoot multiplication in Sorbus commixta Hedl. [58].
The juvenile phase of most plants inherently has a higher rooting potential than the mature phase [27]. Shoots from juvenile seedlings had a higher rooting ability than those from mature material in Diospyros kaki L. [63]. Physiological status also played a significant role in the rooting ability of Sorbus species [34,37,41,43].
In vitro culture can conduce rejuvenation (or reinvigoration), but not always, and it can depend on the number of subcultures in some species. In vitro serial subculture improved rooting in Diospyros kaki [63], Eucalyptus urophylla S. T. Blake [64], apple and cherry rootstocks [65].
Root formation in microcuttings depends on the plant species (genotype) and the cultivation method applied. Even within a species, the rooting ability of microcuttings varies among different cultivars and clones [66]. The effect of genotype was also observed in four Sorbus species; significant differences in rooting ability were found between S. × abscondita, S. gemella, S. × kitaibeliana and S. omissa (Figure 2A). Regardless of the age of the donor plants, Sorbus × abscondita rooted best even when a donor tree was about 50 years old, while S. gemella had the lowest rooting ability of all Sorbus species even when the in vitro culture was established from a 10–15-year-old tree. The opposite situation was found in Sorbus × kitaibeliana, where rooting ability was low (an approximately 80-year-old tree), while S. omissa had a high rooting ability (a 10-year-old tree). These results suggest that genotype plays a large role in rooting ability. Sorbus × abscondita is a member of S. subgen. Soraria Májovský and Bernátová [67] originating from the hybridization of some members of S. subgen. Aria and S. aucuparia. In contrast, S. gemella, S. × kitaibeliana and S. omissa come from the S. subgen. Tormaria Májovský and Bernátová [67], where instead of S. aucuparia, the second parent is S. torminalis. Since the two parent species are quite different ecologically and genetically, the newly created species can show significantly different characteristics accordingly.
One positive aspect of in vitro cultures is that woody species that do not root easily via conventional propagation methods can easily be rooted as microcuttings [68,69,70]. In general, the problems of rooting are more pronounced with woody species [71]. Physiological status played a significant role in the rooting ability of some Sorbus species. Microcuttings derived from juvenile material had a higher rooting ability than those from mature material [34,37,41].
The rooting of microcuttings can take place either in vitro or ex vitro. Rooting in ex vitro environments has two main benefits: it is economic and functional. Rooting under non-sterile conditions should reduce the cost of plant production by microcutting the steps of aseptic manipulation [44]. In vitro rooting of micropropagated plants is an expensive process that can double the final price of these plants [72].
The formation of roots in a tissue culture environment, especially under high humidity and in the presence of sugar, results in morphological and physiological differences compared to roots that develop under normal conditions [73,74,75]. Therefore, in vitro roots must adapt in non-sterile conditions to function properly. By contrast, ex vitro rooted plantlets did not require any additional acclimatization prior to transplanting to regular greenhouse conditions [76].
Ex vitro rooting includes two basic methods to root microcuttings. Both the induction and expression of roots are performed either ex vitro in a greenhouse medium, or they take place in agar or a liquid culture within an ex vitro environment [27].
In vitro rooting of microcuttings has been reported for many Sorbus species and their infrageneric hybrids: S. domestica [34,37], S. aucuparia [43], S. torminalis [30], S. commixta [58], × Malosorbus florentina (Zuccagni) Browicz [41] and also seven rare and endemic Sorbus species [42]. By contrast, ex vitro rooting has been reported only from an experiment with S. torminalis, where root induction took place in vitro and expression occurred under greenhouse conditions [77].
In previous experiments, we tested the rooting potential of Sorbus × abscondita and S. omissa on a half-strength MS agar medium containing IBA or NAA (α-naphthalene acetic acid) at concentrations of 0, 0.5 and 3.0 mg·L–1. The highest rooting (55.6%) was observed in S. × abscondita at 3 mg·L−1 of IBA. In S. omissa, no roots were formed, regardless of the type of auxin and its concentration [78]. When these genotypes were rooted ex vitro, there was a significant increase in rooting ability in S. × abscondita (74%) and in S. omissa (62%—Figure 2A). Similar results were observed in mulberry [68].
Auxin is a key growth regulator in adventitious root formation in plants. Many plants require the presence of auxin for efficient root regeneration [79]. Some of the most important factors affecting in vitro adventitious rooting are the choice of auxin, its concentration and the duration of tissue exposure [80]. IBA is the most often-used agent for rooting microcuttings of a wide variety of species [27]. When microcuttings are rooted in vitro, not only does the level and the duration of auxin treatment differ from ex vitro rooting, but so does the rate of gas exchange by the section of the stem in which the new roots are formed. The presence of auxin in the rooting medium increases the synthesis of ethylene and can have a negative effect on root regeneration [81,82].
The use of various auxins for in vitro root formation in Sorbus species has been reported as follows: Rhizogenesis was induced with IBA in S. commixta and in S. × abscondita [58,79]; in S. domestica, a high percentage of rooting was achieved with both IBA and NAA. A combination of IBA with NAA proved effective in S. torminalis and S. aucuparia [32,43], and a combination of IBA with IAA was found to have an effect in infrageneric hybrid × Malosorbus florentina [41] and NAA has been used with success in seven rare endemic species [42].
In our experiments, we tested the possibility of ex vitro rooting (performing both induction and expression) in four Sorbus species. Rooting was achieved in all the species tested. Rooting ability was high in S. × abscondita and S. omissa but lower in S. gemella and S. × kitaibeliana (Figure 2A). Regarding the effect of auxin treatment on rooting ability, very interesting results were achieved: in S. gemella, S. × kitaibeliana and S. omissa, better results were obtained in rooting without auxin treatment, excluding S. × abscondita, where the highest rooting percentage was observed in the 2% IBA treatment (Figure 2G). A reduction in rooting in the presence of auxin in an agar medium compared to the control (no auxin) has been reported for Anemone L. [83]. The balance of endogenous growth hormones is a key factor in successful organogenesis in plant tissue cultures. Hormonal levels are significantly affected by the addition of exogenous PGRs to the culture medium [84]. The application of PGRs during multiplication can have a negative effect on subsequent rhizogenesis, which has been demonstrated, for example, in S. torminalis [30].
Another factor that can influence rooting in woody plants is the size of the cutting/microcutting [85,86]. Our results showed that determining the optimal length of microcuttings in Sorbus species had a positive effect on rooting. In general, shorter shoots were thinner than longer shoots. The leaves were also less mature. In S. × abscondita, the biggest differences were observed visually in the maturity of the leaves; in addition, shorter shoots were more sensitive to dying.
In conclusion, a successful micropropagation protocol for four Sorbus species was developed. The most difficult phase of the protocol was root induction. The results showed that Sorbus species can be rooted non-sterile in a peat substrate without the presence of auxin. Compared to the other species, Sorbus × abscondita had the highest regeneration and rooting ability. The significantly better results we achieved may have been caused by the influence of a different parental combination of the mentioned species. This protocol will be used in future studies to evaluate the regeneration potential in other species of Sorbus subgen. Tormaria and Sorbus subgen. Soraria [67].
4. Materials and Methods
4.1. Plant Material and In Vitro Culture Initiation
Characteristics of Sorbus species used in our study (Figure 5) and their localities are as follows:
Sorbus × abscondita is a rare hybrid between S. aucuparia and S. danubialis [14] that is treated as a member of S. subgen. Soraria [67]. To this day, it has been recorded at eleven localities in Bohemia (historical part of Czechia) [14], but five of them have gone extinct. Except for two populations, it grows only individually at each locality. The coordinates of the localities of living plants of this taxon are as follows: population in Kladno-Švermov town (Kladno district)—50°10′13.0″ N, 14°07′32.0″ E; population on Sedlo hill near the town of Úštěk (Litoměřice distict)—50°35′31.0″ N, 14°15′43.0″ E; Pochvalovská stráň slope near the village of Pochvalov (Rakovník district)—50°14′11.0″ N, 13°49′03.1″ E; Solopysky village (Louny district)—50°16′08.5″ N, 13°44′25.7″ E; Stříbrník hill near the village of Úholičky (Praha-západ district)—50°10′02.6″ N, 14°21′03.4″ E; Trmice town (Ústí and Labem district)—50°38′07.8″ N, 14°00′44.3″ E.
Sorbus gemella is a triploid species [87] regarded as a member of S. subgen. Tormaria [67], with the supposed parentage of S. danubialis and S. torminalis. Its distribution is limited to the Džbán tableland hills located mostly in the Louny district in the north-central part of Bohemia [14]. The species inhabits edges of marlite plateaus and upper parts of steep slopes on the same bedrock predominantly in oak woodlands. Several subpopulations consist of approximately 1500 individuals. The approximate GPS coordinates of the type locality in Konětopy are 50°16′5.4″ N, 13°44′22.3″ E.
Sorbus × kitaibeliana is a hybrid with the supposed parentage of S. danubialis and S. torminalis (a member of S. subgen. Tormaria). Its only known locality in Czechia is in northern Bohemia near the town of Trmice (Ústí and Labem district—50°38′25″ N, 14°01′09″ E) [14]. There is one known adult, a 12 m high tree, and one 1.5 m high seedling.
Sorbus omissa is another triploid species [88] of the same subgenus and supposed parentage. It is a stenoendemic species occurring in central Bohemia, in the vicinity of the towns of Roztoky and Libčice nad Vltavou (Praha-západ district) in the valley of the lower reach of the river Vltava, where it grows primarily in oak woodlands on humus soils on a substratum of Proterozoic slate. Its two known populations consist of approximately 150 individuals [14,88]. The GPS coordinates of the type specimen of the species are 50°10′06.9″ N, 14°21′09.5″ E.
Figure 5.
Map showing a distribution of Sorbus taxa in Czechia except for Sorbus aucuparia (it is widespread throughout the country). The areas of species with larger distribution were delineated with the “Sample by Buffered Local Adaptive Convex-Hull” tool [89], which combines the creation of a wrapper zone and a minimal convex polygon. Data on Sorbus species with larger distribution were derived from the Pladias database [14,90].
Figure 5.
Map showing a distribution of Sorbus taxa in Czechia except for Sorbus aucuparia (it is widespread throughout the country). The areas of species with larger distribution were delineated with the “Sample by Buffered Local Adaptive Convex-Hull” tool [89], which combines the creation of a wrapper zone and a minimal convex polygon. Data on Sorbus species with larger distribution were derived from the Pladias database [14,90].

All tested plants were triploids (2n = 3x = 51). The DNA ploidy level was estimated using flow cytometry [5]. Carex acutiformis Ehrh. was used as an internal standard.
Plant material of the four Sorbus species, with one clone from each of the selected samples (S. × abscondita—Kladno-Švermov, an approximately 50-year-old donor tree; S. gemella—Konětopy, an approximately 10–15-year-old tree; S. × kitaibeliana—Trmice, an approximately 80-year-old tree and S. omissa—Roztoky, an approximately 10-year-old tree; Figure 6), was identified and collected from natural localities in April or May. Shoots were taken from the lower part of the crown; epicormic sprouts were not available. To obtain in vitro cultures, stem cuttings were taken (40–60 mm in length, Figure 7A,B) and surface-sterilized in 2.5% sodium hypochlorite solution (50% Savo®, Bochemie a.s., Bohumín, The Czech Republic) for 15 min with one drop of Tween-20. Finally, the segments were washed with sterile distilled water three times for 10 min each. Cultures were initiated from apical stem segments with one apical bud and one pair of axillary buds (10–15 mm in length, Figure 7C) and transferred onto a full-strength solid Murashige and Skoog (MS) basal medium with vitamins [91] (Duchefa Biochemie B.V., RV Haarlem, The Netherlands) supplemented with 0.5 mg·L−1 BA (N6-benzyladenine), 0.1 mg·L−1 IBA (indole-3-butyric acid), 20 g·L−1 sucrose, 2 mL·L−1 PPM (Plant Preservative Mixture, Washington, DC, USA) and 7 g·L−1 agar (Sigma-Aldrich Inc., St. Louis, MO, USA). The pH was adjusted to 5.7 using NaOH prior to autoclaving. Explants were cultured in 100 mL Erlenmeyer flasks containing 25 mL of a MS medium. The cultures were maintained in a growth room under a 16 h photoperiod with a photosynthetic photon flux density (PPFD) of 60 µmol·m−2·s provided by cool-white fluorescent tubes (Tungsram, General Electric Company, Boston, MA, USA) at 22 ± 1 °C. Newly grown shoots were divided into two or three parts by subculturing to a fresh medium every three to four weeks. Multiplication and rooting experiments were performed on in vitro cultures that were at least one year old (Figure 8).
4.2. In Vitro Shoot Proliferation
We evaluated the effect of two cytokinins (BA, mT) alone at a concentration of 0.5 mg·L−1 and in combination with an auxin (IBA) at a concentration of 0.1 mg·L−1, and of IBA alone at the same concentration on the induction of shoot formation in four Sorbus species. The plant growth regulator-free medium (PGR) was used as a control. Single shoots (2–3 expanded leaves, ≥1.5 cm long) were excised from the stock cultures and multiplied on an MS-medium containing vitamins, 20 g·L−1 sucrose and 7 g·L−1 agar. Each treatment was repeated three times, with twelve explants per treatment. The cultivation conditions for this experiment were the same as above. The mean number of shoots per explant and total shoot length were recorded after five weeks.
4.3. Ex Vitro Rooting and Acclimatization
Shoots of the four Sorbus species were harvested from an 8–10-week-old in vitro culture maintained on a basal MS medium with 0.5 mg·L−1 BA and 0.1 mg·L−1 IBA, but without PPM. For rooting experiments, microcuttings of two different shoot lengths, 15–25 mm or 26–35 mm, were used. The microcuttings were treated with rooting powder (Rhizopon®AA, Rhizopon BV, Rijndijk, The Netherlands) at IBA concentrations of 1% or 2%.
The microcuttings were cut at the base with a sharp scalpel and immersed in 0.15% antifungal Previcur Energy (Bayer S.A.S., Lyon, France) for 1 min before the application of the rooting powder product (excluding the control). They were then inserted into a plastic dish (14 × 8.5 × 5 cm) with four holes for drainage of excess water, containing a steamed peat-perlite substrate (1:1, v/v) and watered with tap water. Each dish contained eighteen microcuttings. Three dishes were placed in a Minipa plastic box covered with a clear plastic cover with ventilation (tall lid model, Fima, Brno, The Czech Republic) and transferred to a growth room lit with cool-white fluorescent tubes (Tungsram, General Electric Company, Boston, MA, USA) and a photosynthetic photon flux density of 60 µmol·m−2·s−1 for a 16 h photoperiod at 24/19 1 °C (day/night). Each treatment was repeated at least three times, with eighteen microcuttings, i.e., a total of 54 microcuttings per treatment. Root induction (%) and the mean number of roots per microcutting were recorded after six weeks. Then, the rooted plants were replanted into pots (Teku® 10 cm diameter) with a peat substrate Remix-D (Rékyva, Siauliai, Lithuania) and transferred to a greenhouse at 23 °C under natural photoperiod conditions.
4.4. Statistical Analysis
Two-factor analysis of variance and Tukey’s multiple comparison method were used to evaluate differences between the explants. The factors in this case were the Sorbus species and the type of treatment. The dependent variables were the mean number of shoots per explant and the total length of all shoots in the explant. The mean number of shoots per explant could be considered a variable with a Poisson distribution, so the following transformation:
was applied to it [92]. The total length of all explant shoots could be considered a variable with a normal distribution with sufficient accuracy, so its transformation was not necessary.
A three-factor analysis of variance (with the factors Sorbus species, treatment variant and microcutting size) followed by Tukey’s multiple comparison method was used to assess differences between microcuttings. For some dependent variables, the assumption of normal distribution was violated, and various transformations were applied. Specifically, the proportion of rooted microcuttings was a variable with a binomial distribution, so the following transformation:
was applied to it [92]. Again, the mean number of shoots per microcutting could be considered a variable with Poisson distribution, so it was transformed using (1). The results are presented using homogeneous groups at the 0.05 level of significance.
Author Contributions
Conceptualization, J.Š.; methodology, J.Š.; formal analysis, J.Š. and J.V.; investigation, J.Š.; data curation and statistical analysis, J.Š. and D.Z.; writing—original draft preparation, J.Š., J.V. and D.Z.; visualization, J.Š. and J.V.; writing—review and editing, J.Š., J.V. and D.Z.; project administration, J.Š. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Silva Tarouca Research Institute for Landscape and Ornamental Gardening, Public Research Institute (the institutional support VUKOZ-IP-00027073) and by The Technology Agency of the Czech Republic, for the project entitled ‘Conservation methods of the unique Czech gene pool of whitebeams (the genus Sorbus)’, grant number TH03030037.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used in this work are new and original and are fully reported in the present manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Meyer, N.; Meierott, L.; Schuwerk, H.; Angerer, O. Beiträge zur Gattung Sorbus in Bayern. Ber. Bayer. Bot. Ges. Sonderband 2005, 5–216. [Google Scholar]
- Rich, T.C.G.; Houston, L.; Robertson, A.; Proctor, M.C.F. Whitebeams, Rowans and Service Trees of Britain and Ireland. A Monograph of British and Irish Sorbus L.; B.S.B.I. Handbook No. 14.; Botanical Society of the British Isles: London, UK, 2010; pp. 1–223. [Google Scholar]
- Grundt, H.H.; Salvesen, P.H. Kjenn din Sorbus. Rogn og asal i Norge. Rapp. Fra Skog Og Landsk. 2011, 23, 1–104. [Google Scholar]
- Lepší, M.; Lepší, P.; Koutecký, P.; Bílá, J.; Vít, P. Taxonomic revision of Sorbus subgenus Aria occurring in the Czech Republic. Preslia 2015, 87, 109–162. [Google Scholar]
- Velebil, J.; Lepší, M.; Nosková, J.; Lepší, P. Taxonomic assessment of Sorbus subgenus Aria in the Malé Karpaty Mountains. Preslia 2022, 94, 305–334. [Google Scholar] [CrossRef]
- Sennikov, A.N.; Kurtto, A. A phylogenetic checklist of Sorbus s.l. (Rosaceae) in Europe. Memo. Soc. Fauna Fl. Fenn. 2017, 93, 1–78. [Google Scholar]
- Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W. (Eds.) The Global Flora. A Practical Flora to Vascular Plant Species of the World; Special Edition, GLOVAP Nomenclature Part 1, Volume 4; Plant Gateway Ltd.: Bradford, UK, 2018; pp. 1–155. [Google Scholar]
- Kurtto, A.; Sennikov, A.N.; Lampinen, R. (Eds.) Atlas Florae Europaeae. Distribution of Vascular Plants in Europe. 17; Rosaceae (Sorbus s. lato); The Committee for Mapping the Flora of Europe & Societas Biologica Fennica Vanamo: Helsinki, Finland, 2018; pp. 1–132. [Google Scholar]
- Levin, J.; Fay, M.F.; Pellicer, J.; Hedrén, M. Multiple independent origins of intermediate species between Sorbus aucuparia and S. hybrida (Rosaceae) in the Baltic region. Nord. J. Bot. 2018, 36, 1–19. [Google Scholar] [CrossRef]
- Raimondo, F.M.; Gabrieljan, E.; Greuter, W. The genus Aria (Sorbus s. l., Rosaceae) in the Sicilian flora: Taxonomic updating, re-evaluation, description of a new species and two new combinations for one Sicilian and one SW Asian species. Bot. Chron. 2019, 22, 15–37. [Google Scholar]
- Meyer, N.; Feulner, M.; Voss, R.; Rich, T.; Gregor, T.; Paule, J. Sorbus dubronensis, eine neue endemische Art aus der Untergattung Aria (Sorbus s. l., Rosaceae) für Süddeutschland, und ihre Abgrenzung zu verwandten Arten. Ber. Bayer. Bot. Ges. 2020, 90, 83–106. [Google Scholar]
- Meyer, N.; Meierott, L. Ergänzende Beiträge zur Sorbus-Flora von Bayern. Ber. Bayer. Bot. Ges. 2021, 91, 21–48. [Google Scholar]
- Rivers, M.C.; Beech, E.; Bazos, I.; Bogunić, F.; Buira, A.; Caković, D.; Carapeto, A.; Carta, A.; Cornier, B.; Fenu, G.; et al. European Red List of Trees; IUCN: Cambridge, UK; Brussels, Belgium, 2019; pp. 1–59.
- Kaplan, Z.; Danihelka, J.; Lepší, M.; Lepší, P.; Ekrt, L.; Chrtek, J., Jr.; Kocián, J.; Prančl, J.; Kobrlová, L.; Hroneš, M.; et al. Distributions of vascular plants in the Czech Republic. Part 3. Preslia 2016, 88, 459–544. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Chrtek, J., Jr.; Kirschner, J.; Kubát, K.; Štech, M.; Štěpánek, J. (Eds.) Klíč ke Květeně České Republiky, 2nd ed.; Academia: Praha, Czech Republic, 2019; pp. 1–1168. (In Czech) [Google Scholar]
- Grulich, V. Červený seznam cévnatých rostlin ČR. Příroda 2017, 35, 75–132. (In Czech) [Google Scholar]
- Kociánová, M.; Štursová, H. Revize rozšíření a ekologie jeřábu krkonošského (Sorbus sudetica). Opera Corcon. 1986, 23, 77–110. (In Czech) [Google Scholar]
- Kovanda, M. Jeřáby (Sorbus) České republiky a jejich ochrana. Příroda 1999, 15, 31–47. (In Czech) [Google Scholar]
- Kociánová, M.; Štursová, H.; Zahradníková, J. Klonální růst endemického jeřábu Sorbus sudetica v Krkonoších. Opera Corcon. 2005, 42, 99–105. (In Czech) [Google Scholar]
- Pías, B.; Guitián, P. Breeding system and pollen limitation in the masting tree Sorbus aucuparia L. (Rosaceae) in the NW Iberian Peninsula. Acta Oecol. 2006, 29, 97–103. [Google Scholar] [CrossRef]
- Hoebee, S.E.; Arnold, U.; Düggelin, C.; Gugerli, F.; Brodbeck, S.; Rotach, P.; Holderegger, R. Mating patterns and contemporary gene flow by pollen in a large continuous and a small isolated population of the scattered forest tree Sorbus torminalis. Heredity 2007, 99, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Liljefors, A. Studies on propagation, embryology, and pollination in Sorbus. Acta Horti Bergiani 1953, 16, 277–329. [Google Scholar]
- Jankun, A.; Kovanda, M. Apomixis in Sorbus sudetica (Embryological studies in Sorbus 1). Preslia 1986, 58, 7–19. [Google Scholar]
- Jankun, A.; Kovanda, M. Apomixis and origin of Sorbus bohemica (Embryological studies in Sorbus 2). Preslia 1987, 59, 97–116. [Google Scholar]
- Dickinson, T.A.; Campbell, C.S. Population Structure and Reproductive Ecology in the Maloideae (Rosaceae). Syst. Bot. 1991, 16, 350–362. [Google Scholar] [CrossRef]
- Bärtels, A. Rozmnožování Dřevin; Státní Pedagogické Nakladatelství: Praha, Czechoslovakia, 1988; pp. 1–452. (In Czech) [Google Scholar]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. Hartmann and Kester’s Plant Propagation: Principles and Practices, 8th ed.; Prentice Hall: Hoboken, NJ, USA, 2011; pp. 1–915. [Google Scholar]
- Dujíčková, M.; Malá, J.; Chalupa, V. Vegetativní množení Sorbus torminalis (L.) Crantz a Sorbus domestica L. in vitro. Práce Vúlhm 1991, 77, 27–48. (In Czech) [Google Scholar]
- Malá, J.; Máchová, P.; Cvrčková, H.; Čížková, L. Využití mikropropagace pro reprodukci genových zdrojů vybraných ušlechtilých listnatých dřevin (Malus sylvestris, Pyrus pyraster, Sorbus torminalis, S. aucuparia a Prunus avium). Zprávy Lesn. Výzkumu 2005, 4, 219–224. (In Czech) [Google Scholar]
- Malá, J.; Máchová, P.; Cvrčková, H.; Karady, M.; Novák, O.; Mikulík, J.; Hauserová, E.; Greplová, J.; Strnad, M.; Doležal, K. Micropropagation of wild service tree (Sorbus torminalis (L.) Crantz): The regulative role of different aromatic cytokinins during organogenesis. J. Plant Growth Regul. 2009, 28, 341–348. [Google Scholar] [CrossRef]
- Chalupa, V. In vitro propagation of willows (Salix spp.), European mountain-ash (Sorbus aucuparia L.) and black locust (Robinia pseudoacacia L.). Biol. Plant. 1983, 25, 305–307. [Google Scholar] [CrossRef]
- Chalupa, V. Vegetativní rozmnožování listnatých dřevin řízky a metodou in vitro. Lesnictví 1987, 33, 501–510. (In Czech) [Google Scholar]
- Lall, S.; Mandegaran, Z.; Roberts, A.V. Shoot multiplication and adventitious regeneration in Sorbus aucuparia. Plant Cell Tissue Organ Cult. 2006, 85, 23–29. [Google Scholar] [CrossRef]
- Arrillaga, I.; Marzo, T.; Segura, J. Micropropagation of juvenile and adult Sorbus domestica L. Plant Cell Tissue Organ Cult. 1991, 27, 341–348. [Google Scholar] [CrossRef]
- Meier-Dinkel, A. In vitro Vermehrung von Speierling (Sorbus domestica L.). Corminaria 1998, 9, 9–13. [Google Scholar]
- Miko, M.; Gažo, J.; Biroščíková, M. In vitro klonové množenie genetických zdrojov jarabiny oskorušovej (Sorbus domestica L.) z územia Slovenska. Acta Fytotechn. Zootechn. 2004, 7, 85–89. (In Slovak) [Google Scholar]
- Nikolaou, P.; Zagas, D.; Scaltsoyiannes, V.; Balas, E.; Xilogianni, V.; Tsoulpha, P.; Tsaktsira, M.; Voulgaridou, E.; Iliev, I.; Triantafyllou, K.; et al. Advances in the micropropagation of service tree (Sorbus domestica L.). Propag. Ornam. Plants 2008, 8, 154–157. [Google Scholar]
- Ďurkovič, J.; Mišalová, A. Wood formation during ex vitro acclimatisation in micropropagated true service tree (Sorbus domestica L.). Plant Cell Tissue Organ Cult. 2009, 96, 343–348. [Google Scholar] [CrossRef]
- Malá, J.; Cvrčková, H.; Máchová, P.; Dostál, J. Mikropropagace Jeřábu Oskeruše (Sorbus domestica L.); Certified Methodology 4/2011; Forestry and Game Management Research Institute: Strnady, Czech Republic, 2011; pp. 1–17. (In Czech) [Google Scholar]
- Piagnani, M.C.; Zaccheo, P.; Crippa, L. Micropropagation of service tree (Sorbus domestica L.): Role of some factors on in vitro proliferation and rooting, and extra vitro acclimatization. Agrochimica 2012, 56, 219–233. [Google Scholar]
- Martini, A.N.; Papafotiou, M. Season and explant origin affect phenolic content, browning of explants, and micropropagation of ×Malosorbus florentina (Zucc.) Browicz. HortScience 2013, 48, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Máchová, P.; Malá, J.; Cvrčková, H.; Dostál, J.; Buriánek, V. In vitro reproduction of rare and endemic species of rowan tree. J. For. Sci. 2013, 59, 386–390. [Google Scholar] [CrossRef] [Green Version]
- Chalupa, V. In vitro propagation of mature trees of Sorbus aucuparia L. and field performance of micropropagated tree. J. For. Sci. 2002, 48, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Bhojwani, S.S.; Razdan, M.K. Plant Tissue Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 1983; pp. 1–502. [Google Scholar]
- Hackett, W.P. Juvenility, maturation and rejuvenation in woody plants. Hort. Rev. 1985, 7, 109–155. [Google Scholar]
- Basheer-Salimia, R. Juvenility, maturity and rejuvenation in woody plants. Hebron Univ. Res. J. 2007, 3, 17–43. [Google Scholar]
- Ďurkovič, J.; Mišalová, A. Micropropagation of temperate noble hardwoods: An overview. Funct. Plant Sci. Biotechnol. 2008, 2, 1–19. [Google Scholar]
- Werbrouck, S.P.O.; van der Jeugt, B.; Dewitte, W.; Prinsen, E.; Van Onckelen, H.A.; Debergh, P.C. The metabolism of benzyladenine in Spathiphyllum floribundum ‘Schott Petite’ in relation to acclimatisation problems. Plant Cell Tissue Organ Cult. 1995, 14, 662–665. [Google Scholar] [CrossRef]
- Bairu, M.W.; Stirk, W.A.; Doležal, K.; Van Staden, J. Optimizing the micropropagation protocol for the endangered Aloe polyphylla: Can meta-topolin and its derivatives serve as replacement for benzyladenine and zeatin? Plant Cell Tissue Organ Cult. 2007, 90, 15–23. [Google Scholar] [CrossRef]
- Tarkowská, D.; Doležal, K.; Tarkowski, P.; Astot, C.; Holub, J.; Fuksová, K.; Schmülling, T.; Sandberg, G.; Strnad, M. Identification of new aromatic cytokinins in Arabidopsis thaliana and Populus × canadensis leaves by LC-(+)ESI-MS and capillary liquid chromatography/frit-fast atom bombardment mass spektrometry. Physiol. Plant. 2003, 117, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Strnad, M.; Hanuš, J.; Vaněk, T.; Kamínek, M.; Ballantine, J.A.; Fussell, B.; Hanke, D.E. Meta-topolin, a highly active aromatic cytokinin from poplar leaves (Populus × canadensis Moench., cv. Robusta. Phytochemistry 1997, 45, 213–218. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; Strnad, M.; Van Onckelen, H.A.; Debergh, P.C. Meta-topolin, an alternative to benzyladenine in tissue culture? Physiol. Pl. 2008, 98, 291–297. [Google Scholar] [CrossRef]
- Amoo, S.O.; Van Staden, J. Influence of plant growth regulators on shoot proliferation and secondary metabolite production in micpropagated Huernia hystrix. Plant Cell Tissue Organ Cult. 2013, 112, 249–256. [Google Scholar] [CrossRef]
- Moyo, M.; Finnie, J.F.; Van Staden, J. Recalcitrant effects associated with the development of basal callus-like tissue on caulogenesis and rhizogenesis in Sclerocarya birrea. Plant Growth Regul. 2011, 63, 187–195. [Google Scholar] [CrossRef]
- Šedivá, J.; Vlašínová, H.; Klemš, M.; Vejsadová, H.; Švajdlenka, E.; Mertelík, J.; Kloudová, K.; Havel, L. Behaviour of resistant and non-resistant clones of Aesculus hippocastanum in vitro. Acta Hort. 2013, 988, 123–128. [Google Scholar] [CrossRef]
- van der Westhuizen, A. The use of meta-topolin as an alternative cytokinin in the tissue culture of Eucalyptus species. Acta Hort. 2014, 1055, 25–28. [Google Scholar] [CrossRef]
- Ördögh, M.; Jambor-Benczúr, E.; Tilly-Mándy, A.; Lelik, L. Effects of different cytokinins on proliferation of Sorbus borbasii ‘Herkulesfürdö’. Propag. Ornam. Plants 2009, 9, 43–46. [Google Scholar]
- Jeong, B.R.; Sivanesan, I. Micropropagation of Sorbus commixta Hedl. Propag. Ornam. Plants 2015, 15, 142–146. [Google Scholar]
- Ördögh, M.; Jambor-Benczúr, E.; Tilly-Mándy, A.; Lelik, L. The effects of growth regulators in proliferation of Sorbus redliana “Burokvölgy”. Int. J. Hortic. Sci. 2006, 12, 77–83. [Google Scholar] [CrossRef]
- Meyer, E.M.; Touchell, D.H.; Ranney, T.G. In vitro shoot regeneration and polyploidy induction from leaves of Hypericum species. Hortscience 2009, 44, 1957–1961. [Google Scholar] [CrossRef] [Green Version]
- Kucharska, D.; Orlikowska, T.; Maciorowski, R.; Kunka, M.; Wójcik, D.; Pluta, S. Application of meta-Topolin for improving micropropagation of gooseberry (Ribes grossularia). Sci. Hort. 2020, 272, 109529. [Google Scholar] [CrossRef]
- Hlophe, N.P.; Aremu, A.O.; Doležal, K.; Van Staden, J.; Finnie, J.F. Cytokinin-facilitated plant regeneration of three Brachystelma species with different conservation status. Plants 2020, 9, 1657. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Ito, J.; Sugiura, A. Comparison of growth and rooting characteristics of micropropagated adult plants and juvenile seedlings of persimmon (Diospyros kaki L.). J. Jpn. Soc. Hortic. Sci. 1994, 63, 537–541. [Google Scholar] [CrossRef]
- Mendonça, E.G.; Batista, T.R.; Stein, V.C.; Balieiro, F.P.; Abreu, J.R.; Pires, M.F.; de Souza, P.A.; Paiva, L.V. In vitro serial subculture to improve rooting of Eucalyptus urophylla. New For. 2020, 51, 801–816. [Google Scholar] [CrossRef]
- Grant, N.J.; Hammatt, N. Increased root and shoot production during micropropagation of cherry and apple rootstocks: Effect of subculture frequency. Tree Physiol. 1999, 19, 899–903. [Google Scholar] [CrossRef] [Green Version]
- Economou, A.S. From microcutting rooting to microplant establishment: Key points to consider for maximum success in woody plants. Acta Horicult. 2013, 988, 43–56. [Google Scholar] [CrossRef]
- Májovský, J.; Bernátová, D. Nové hybridogénne podrody rodu Sorbus L. emend. Crantz. Acta Hortic. Regiotect. 2001, 1, 20–21. (In Slovak) [Google Scholar]
- Aroonpong, P.; Chang, J.C. Micropropagation of a difficult-to-root weeping mulberry (Morus alba var. Shidareguwa): A popular variety for ornamental purposes. Sci. Hort. 2015, 194, 320–326. [Google Scholar] [CrossRef]
- Iapichino, G.; Airo, M. Multiplication of Crataegus monogyna by in vitro culture of nodal segments. Acta Hort. 2009, 812, 135–140. [Google Scholar] [CrossRef]
- Noshad, D.; Miresmaili, S.; Riseman, A.; Ekramoddoullah, A. In vitro propagation of seven Daphne L. species. Plant Cell Tissue Organ Cult. 2009, 96, 201–209. [Google Scholar] [CrossRef]
- Debergh, P.C.; Topoonyanont, N.; Van Huylenbroeck, J.; Moreira da Silva, H.; Oyaert, E. Preparation of microplants for ex vitro establishment. Acta Hort. 2000, 530, 269–276. [Google Scholar] [CrossRef]
- Herman, E.B. Recent Advances in Plant Tissue Culture XVIII. Plant Micropropagation: 2010–2013; Agritech Consultants, Inc.: Mohegan Lake, NY, USA, 2013; pp. 1–138. [Google Scholar]
- Apter, R.C.; McWilliams, E.L.; Davies, F.T., Jr. In vitro and ex vitro adventitious root formation in Asian jasmine (Trachelospermum asiaticum). I. Comparative morphology. J. Am. Soc. Hort. Sci. 1993, 118, 902–905. [Google Scholar] [CrossRef] [Green Version]
- Soukup, A.; Malá, J.; Hrubcová, M.; Kálal, J.; Votrubová, O.; Cvikrová, M. Differences in anatomical structure and lignin content of roots of pedunculate oak and wild cherry-tree plantlets during acclimation. Biol. Pl. 2004, 48, 481–489. [Google Scholar] [CrossRef]
- Hatzilazarou, S.P.; Syros, T.D.; Yupsanis, T.A.; Bosabalidis, A.M.; Economou, A.S. Peroxidases, lignin and anatomy during in vitro and ex vitro rooting of gardenia (Gardenia jasminoides Ellis) microshoots. J. Pl. Physiol. 2006, 163, 827–836. [Google Scholar] [CrossRef]
- Pruski, K.W.; Tina, L.; Astatkie, T.; Nowak, J. Micropropagation of chokecherry and pincherry cultivars. Plant Cell Tissue Organ Cult. 2000, 63, 93–100. [Google Scholar] [CrossRef]
- Marques, S.L.; Canhoto, J.; Gonçalves, J.C.; Diogo, M.G. Micropropagation of Sorbus torminalis (L.) Crantz: Hormonal effects during multiplication and rooting phases. Acta Hort. 2013, 990, 397–404. [Google Scholar] [CrossRef]
- Šedivá, J.; Businský, R.; Pospíšková, M.; Velebil, J.; Drahošová, H.; Zýka, V. Conservation methods of Czech gene pool of whitebeams. Acta Hort. 2021, 1324, 41–46. [Google Scholar] [CrossRef]
- Pierik, R.L.M. In Vitro Culture of Higher Plants; Kluwer Academic Publishers Group: Dordrecht, The Netherlands, 1989; pp. 1–344. [Google Scholar]
- De Klerk, G.J.; Van der Krieken, W.; De Jong, J.C. Review the formation of adventitious roots: New concepts, new possibilities. Vitr. Cell Dev. Biol.-Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]
- De Klerk, G.J.M. Rooting in vitro. Unexpected effects of deficient gas exchange. Prophyta Annu. 2013, 2013, 46–47. [Google Scholar]
- De Klerk, G.J.M.; Paffen, A.K. MnO4-coated grains prevent a massive, detrimental rise of ethylene during in vitro rooting of rose microcuttings. Propag. Ornam. Plants 2020, 20, 96–102. [Google Scholar]
- Šedivá, J.; Zahumenická, P.; Fernández Cusimamani, E. An efficient in vitro propagation protocol for snowdrop anemone (Anemone sylvestris L.). Hort. Sci. 2017, 44, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Sreekissoon, A.; Plačková, L.; Doležal, K.; Finnie, J.F.; Van Staden, J. In vitro and ex vitro vegetative propagation and cytokinin profiles of Sceletium tortuosum (L.) N. E. Br.: A South African medicinal plant. Plant Cell Tissue Organ Cult. 2021, 145, 191–202. [Google Scholar] [CrossRef]
- Haq, I.U.; Ahmad, T.; Hafiz, I.A.; Abbasi, N.A. Influence of microcutting sizes and IBA concentrations on in vitro rooting of olive cv. ‘Dolce Agogia’. Pak. J. Bot. 2009, 41, 1213–1222. [Google Scholar]
- OuYang, F.; Wang, J.; Li, Y. Effects of cutting size and exogenous hormone treatment on rooting of shoot cuttings in Norway spruce [Picea abies (L.) Karst.]. New For. 2015, 46, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Lepší, M.; Koutecký, P.; Nosková, J.; Lepší, P.; Urfus, T.; Rich, T.C.G. Versatility of reproductive modes and ploidy level interactions in Sorbus s.l. (Malinae, Rosaceae). Bot. J. Linn. Soc. 2019, 191, 502–522. [Google Scholar] [CrossRef]
- Velebil, J. Sorbus omissa, a new endemic hybridogenous species from the lower Vltava river valley. Preslia 2012, 84, 375–390. [Google Scholar]
- Sdmtoolbox. Available online: http://www.sdmtoolbox.org (accessed on 7 July 2022).
- Pladias. Database of the Czech Flora and Vegetation. Available online: http://www.pladias.cz/en/ (accessed on 7 July 2022).
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Montgomery, D.C. Design and Analysis of Experiments; Willey: New York, NY, USA, 2012; pp. 1–730. [Google Scholar]
Figure 1.
Shoot production in Sorbus species cultured with different combinations of N6-benzyladenine (BA), indole-3-butyric acid (IBA) and meta-topolin (mT), applied at 0.5 mg·L−1, 0.1 mg·L−1 and 0.5 mg·L−1, respectively. (A,B) The effect of genotype on the mean number of shoots per explant (A) and the total shoot length (B). (C,D) The effect of plant growth regulator (PGR) treatment on the mean number of shoots per explant (C) and on the total shoot length (D). Values are mean standard errors. In each graph, different letter(s) on the bars show significant differences according to Tukey’s range test (p-value = 0.05). Sorbus × abscondita (SA), S. gemella (SG), S. × kitaibeliana (SK), S. omissa (SO).
Figure 1.
Shoot production in Sorbus species cultured with different combinations of N6-benzyladenine (BA), indole-3-butyric acid (IBA) and meta-topolin (mT), applied at 0.5 mg·L−1, 0.1 mg·L−1 and 0.5 mg·L−1, respectively. (A,B) The effect of genotype on the mean number of shoots per explant (A) and the total shoot length (B). (C,D) The effect of plant growth regulator (PGR) treatment on the mean number of shoots per explant (C) and on the total shoot length (D). Values are mean standard errors. In each graph, different letter(s) on the bars show significant differences according to Tukey’s range test (p-value = 0.05). Sorbus × abscondita (SA), S. gemella (SG), S. × kitaibeliana (SK), S. omissa (SO).

Figure 2.
Rooting of microcuttings of Sorbus species treated with different concentrations of indole-3-butyric acid (IBA), applied at 0%, 1% and 2%. (A,B) The effect of genotype on the rooting percentage (A) and the mean number of roots per microcutting (B). (C,D) The effect of auxin treatment on the rooting percentage (C) and the mean number of roots per microcutting (D). (E,F) The effect of the size of the microcutting on the rooting percentage (E) and the mean number of roots per microcutting (F). (G,H) The interaction between the genotype and auxin treatment on the rooting percentage (G) and the mean number of roots per microcutting (H). (I) The interaction between the genotype and the size of the microcutting on the rooting percentage. Values are mean standard errors. In each graph, different letter(s) on the bars show significant differences according to Tukey’s range test (p-value < 0.05). Sorbus × abscondita (SA), S. gemella (SG), S. × kitaibeliana (SK), S. omissa (SO).
Figure 2.
Rooting of microcuttings of Sorbus species treated with different concentrations of indole-3-butyric acid (IBA), applied at 0%, 1% and 2%. (A,B) The effect of genotype on the rooting percentage (A) and the mean number of roots per microcutting (B). (C,D) The effect of auxin treatment on the rooting percentage (C) and the mean number of roots per microcutting (D). (E,F) The effect of the size of the microcutting on the rooting percentage (E) and the mean number of roots per microcutting (F). (G,H) The interaction between the genotype and auxin treatment on the rooting percentage (G) and the mean number of roots per microcutting (H). (I) The interaction between the genotype and the size of the microcutting on the rooting percentage. Values are mean standard errors. In each graph, different letter(s) on the bars show significant differences according to Tukey’s range test (p-value < 0.05). Sorbus × abscondita (SA), S. gemella (SG), S. × kitaibeliana (SK), S. omissa (SO).



Figure 3.
Comparison of the development of microcuttings in Sorbus species in the best rooting treatment in a steamed peat-perlite substrate after six weeks. (A) Sorbus × abscondita, microcuttings treated with 2% indole-3-butyric acid (IBA) rooting powder. (B) Sorbus gemella, the control. (C) Sorbus × kitaibeliana, the control. (D) Sorbus omissa, the control. (E) Detail of rooted plantlets of Sorbus × abscondita. (F) Detail of rooted plantlets of Sorbus omissa.
Figure 3.
Comparison of the development of microcuttings in Sorbus species in the best rooting treatment in a steamed peat-perlite substrate after six weeks. (A) Sorbus × abscondita, microcuttings treated with 2% indole-3-butyric acid (IBA) rooting powder. (B) Sorbus gemella, the control. (C) Sorbus × kitaibeliana, the control. (D) Sorbus omissa, the control. (E) Detail of rooted plantlets of Sorbus × abscondita. (F) Detail of rooted plantlets of Sorbus omissa.

Figure 4.
Acclimatization and development of rooted plants of Sorbus species. (A) 1-year-old plants (S. × abscondita and S. omissa). (B) 2-year-old plants (S. × kitaibeliana). (C) 3-year-old plants (S. × abscondita).
Figure 4.
Acclimatization and development of rooted plants of Sorbus species. (A) 1-year-old plants (S. × abscondita and S. omissa). (B) 2-year-old plants (S. × kitaibeliana). (C) 3-year-old plants (S. × abscondita).

Figure 6.
Sorbus species investigated. (A) Sorbus × abscondita. (B) Sorbus gemella. (C) Sorbus × kitaibeliana. (D) Sorbus omissa.
Figure 6.
Sorbus species investigated. (A) Sorbus × abscondita. (B) Sorbus gemella. (C) Sorbus × kitaibeliana. (D) Sorbus omissa.

Figure 7.
Explant source in Sorbus sp. (A) Original shoot. (B) Prepared shoot before explant disinfestation. (C) Initiation explant.
Figure 7.
Explant source in Sorbus sp. (A) Original shoot. (B) Prepared shoot before explant disinfestation. (C) Initiation explant.

Figure 8.
Bulking of stock material in Sorbus species on full-strength solid Murashige and Skoog (MS) basal medium with vitamins supplemented with 0.5 mg·L–1 N6-benzyladenine (BA) and 0.1 mg·L–1 indole-3-butyric acid (IBA). (A) In vitro collection of Sorbus species. (B) Detail of shoot regeneration in Sorbus × abscondita.
Figure 8.
Bulking of stock material in Sorbus species on full-strength solid Murashige and Skoog (MS) basal medium with vitamins supplemented with 0.5 mg·L–1 N6-benzyladenine (BA) and 0.1 mg·L–1 indole-3-butyric acid (IBA). (A) In vitro collection of Sorbus species. (B) Detail of shoot regeneration in Sorbus × abscondita.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The effect of interaction between the genotype and the PGR treatment on the mean number of shoots per explant in Sorbus species (n). The term SE denotes corresponding standard error; group shows significant differences according to Tukey’s range test (p-value = 0.05).
Table 1.
The effect of interaction between the genotype and the PGR treatment on the mean number of shoots per explant in Sorbus species (n). The term SE denotes corresponding standard error; group shows significant differences according to Tukey’s range test (p-value = 0.05).
| PGR | Genotype | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. × abscondita | S. gemella | S. × kitaibeliana | S. omissa | |||||||||
| n | SE | Group | n | SE | Group | n | SE | Group | n | SE | Group | |
| Control | 2.00 | 0.28 | abcd | 1.11 | 0.09 | abcd | 1.28 | 0.09 | ab | 1.00 | 0.00 | a |
| BA | 2.97 | 0.41 | de | 2.36 | 0.39 | bcde | 3.42 | 0.59 | de | 2.42 | 0.36 | abcde |
| BA+IBA | 2.42 | 0.33 | abcde | 1.69 | 0.22 | abcd | 2.42 | 0.33 | abcde | 2.11 | 0.27 | abcd |
| mT | 3.72 | 0.36 | e | 2.39 | 0.42 | abcde | 2.83 | 0.36 | cde | 1.89 | 0.14 | abcd |
| mT+IBA | 3.47 | 0.31 | e | 2.69 | 0.43 | bcde | 2.81 | 0.34 | cde | 2.13 | 0.37 | abcd |
| IBA | 1.69 | 0.33 | abcd | 1.03 | 0.03 | a | 1.36 | 0.12 | abc | 1.03 | 0.03 | a |
Plant growth regulator (PGR), N6-benzyladenine (BA), indole-3-butyric acid (IBA) and meta-topolin (mT).
Table 2.
The effect of interaction between the genotype and the PGR treatment on the total shoot length in Sorbus species (l). The term SE denotes corresponding standard error; group shows significant differences according to Tukey’s range test (p-value = 0.05).
Table 2.
The effect of interaction between the genotype and the PGR treatment on the total shoot length in Sorbus species (l). The term SE denotes corresponding standard error; group shows significant differences according to Tukey’s range test (p-value = 0.05).
| PGR | Genotype | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. × abscondita | S. gemella | S. × kitaibeliana | S. omissa | |||||||||
| l | SE | Group | l | SE | Group | l | SE | Group | l | SE | Group | |
| Control | 56.69 | 8.9 | bcdef | 22.97 | 1.0 | a | 31.69 | 3.6 | abc | 20.67 | 0.6 | a |
| BA | 59.92 | 8.3 | cdef | 40.56 | 5.3 | abcde | 71.00 | 12.9 | ef | 42.36 | 5.7 | abcde |
| BA+IBA | 54.94 | 7.8 | abcde | 30.50 | 3.2 | abc | 56.03 | 7.6 | bcde | 42.03 | 5.2 | abcde |
| mT | 68.13 | 6.5 | def | 50.53 | 5.5 | abcde | 61.81 | 5.9 | cdef | 36.08 | 1.9 | abcd |
| mT+IBA | 89.22 | 8.1 | f | 44.86 | 4.1 | abcde | 65.50 | 6.3 | def | 50.49 | 8.8 | abcde |
| IBA | 39.99 | 7.3 | abcde | 27.00 | 1.2 | ab | 36.31 | 3.9 | abcd | 24.25 | 0.8 | ab |
Plant growth regulator (PGR), N6-benzyladenine (BA), indole-3-butyric acid (IBA) and meta-topolin (mT).
Table 3.
Effect of different factors (genotype, auxin treatment and microcutting size) on rooting of Sorbus species used in our study.
Table 3.
Effect of different factors (genotype, auxin treatment and microcutting size) on rooting of Sorbus species used in our study.
| Rooting | Mean Number of Roots per Microcutting | |||||||
|---|---|---|---|---|---|---|---|---|
| df | Mean Squares | F | df | Mean Squares | F | |||
| Genotype | 3 | 2.42 | 53.98 | *** | 3 | 23.06 | 72.66 | *** |
| Auxin treatment | 2 | 1.13 | 25.31 | *** | 2 | 5.83 | 18.37 | *** |
| Microcutting size | 1 | 1.15 | 25.71 | *** | 1 | 1.52 | 4.80 | * |
| Genotype × auxin treatment | 6 | 0.35 | 7.97 | *** | 6 | 3.98 | 12.56 | *** |
| Genotype × microcutting size | 3 | 0.25 | 5.65 | ** | 3 | 0.48 | 15.14 | ns |
| Auxin treatment × microcutting size | 2 | 0.02 | 0.59 | ns | 2 | 0.10 | 0.32 | ns |
| Error | 54 | 0.04 | 568 | 0.31 | ||||
*, **, ***, ns—Tukey’s range test significant at p = 0.05, p = 0.01, p = 0.001 or not significant; df means degrees of freedom, and F is the F-statistic.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Šedivá, J.; Velebil, J.; Zahradník, D. Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia. Plants 2023, 12, 488. https://doi.org/10.3390/plants12030488
AMA Style
Šedivá J, Velebil J, Zahradník D. Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia. Plants. 2023; 12(3):488. https://doi.org/10.3390/plants12030488
Chicago/Turabian StyleŠedivá, Jana, Jiří Velebil, and Daniel Zahradník. 2023. "Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia" Plants 12, no. 3: 488. https://doi.org/10.3390/plants12030488
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.