
From the Wild to the Field: Documentation, Propagation, Pilot Cultivation, Fertilization, and Phytochemical Evaluation of the Neglected and Underutilized Amelanchier ovalis Medik. (Rosaceae)

 ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Asexual Propagation of the Greek Native Germplasm
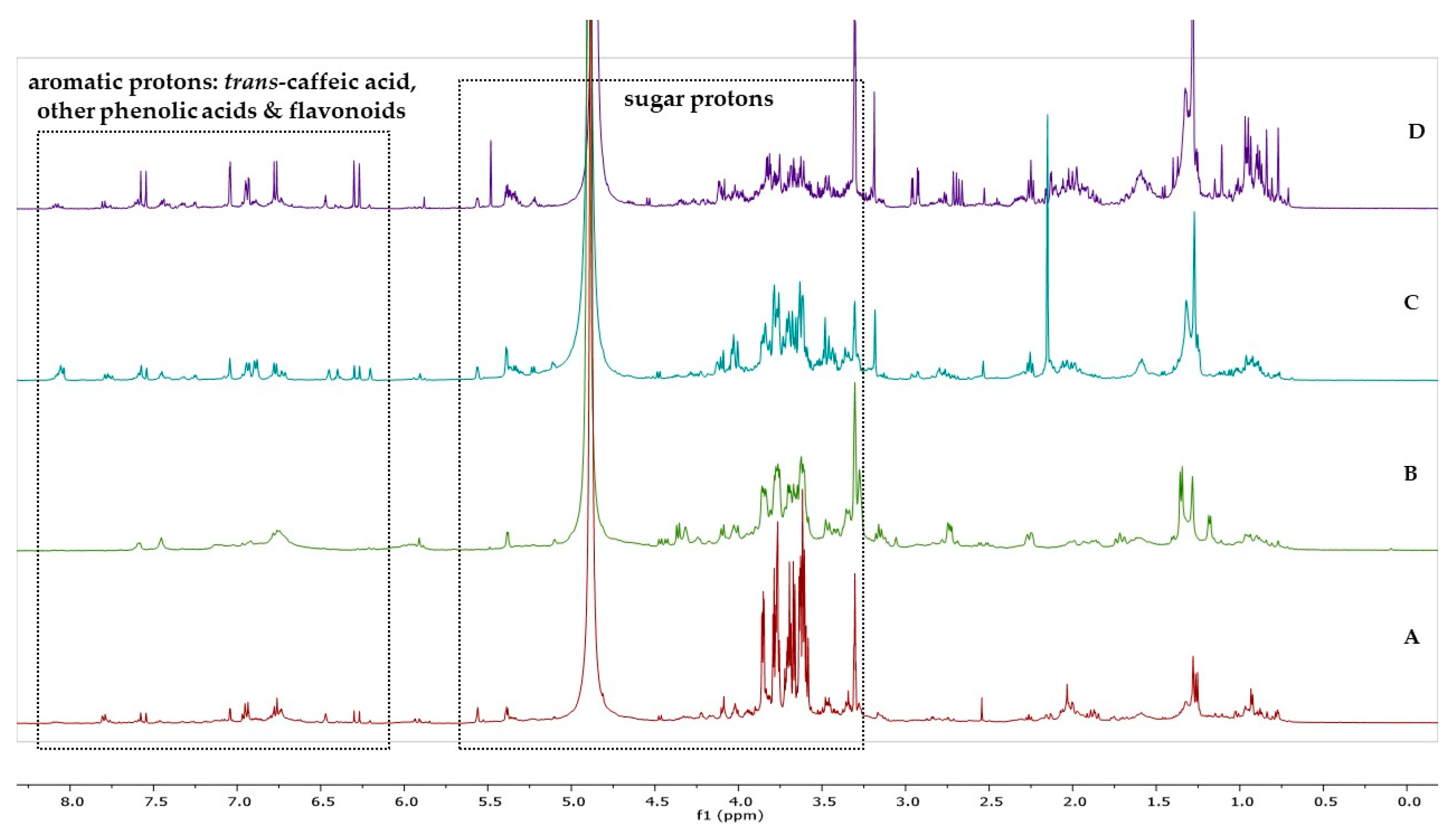
2.2. Total Phenolic Content and Antioxidant Activity
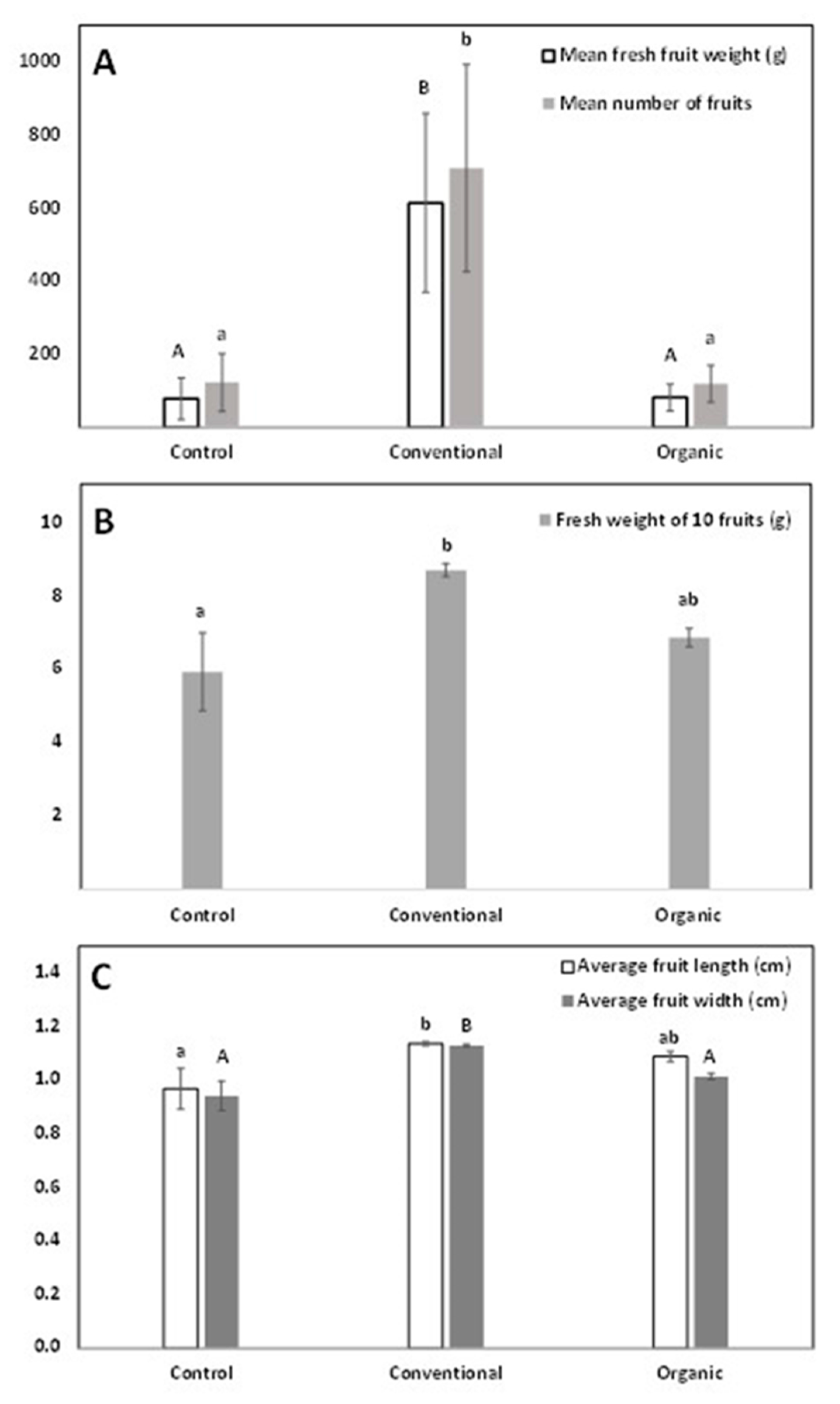
2.3. Field Cultivation Trial under Different Fertilization Regimes
3. Discussion
3.1. Asexual Propagation of Greek Native Amelanchier ovalis
3.2. Total Phenolic Content and Antioxidant Activity of Greek Native Amelanchier Ovalis
3.3. Pilot Orchard-Type Cultivation of Greek Native Amelanchier ovalis
4. Materials and Methods
4.1. Documentation of Plant Material
4.2. Preliminary Propagation Trials of the Collected and Documented Greek Germplasm
4.3. Propagation Experiments with Greek Germplasm
4.4. In Vitro Assessment of the Total Phenolic Content and Antioxidant Activity in Different Organs of the Greek Germplasm
4.4.1. Extracts from Plant Material
4.4.2. Determination of Total Phenolic Content (TPC)
4.4.3. Evaluation of Antioxidant Activity (AA)
4.5. Pivotal Field Cultivation Trial of the Greek Germplasm
4.6. Experimental Design and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Libiad, M.; Khabbach, A.; El Haissoufi, M.; Anestis, I.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Greveniotis, V.; Tsiripidis, I.; et al. Agro-Alimentary potential of the neglected and underutilized local endemic plants of Crete (Greece), Rif-Mediterranean Coast of Morocco and Tunisia: Perspectives and challenges. Plants 2021, 10, 1770. [Google Scholar] [CrossRef] [PubMed]
- Ercisli, S. A short review of the fruit germplasm resources of Turkey. Gen. Res. Crop Evol. 2004, 51, 419–435. [Google Scholar] [CrossRef]
- Verma, N.; Mohanty, A.; Lal, A. Pomegranate genetic resources and germplasm conservation: A review. Fruit Veg. Cereal Sci. Biotechnol. 2010, 4, 120–125. Available online: http://www.globalsciencebooks.info/Online/GSBOnline/images/2010/FVCSB_4(SI2)/FVCSB_4(SI2)120-125o.pdf (accessed on 31 January 2023).
- Botu, M.; Botu, I.; Achim, G.; Preda, S.; Scutelnicu, A.; Giura, S. Conservation of fruit tree genetic resources and their use in the breeding process. Ann. Valahia Univ. Targoviste 2017, 11, 66–69. [Google Scholar] [CrossRef] [Green Version]
- Manco, R.; Basile, B.; Capuozzo, C.; Scognamiglio, P.; Forlani, M.; Rao, R.; Corrado, G. Molecular and phenotypic diversity of traditional European plum (Prunus domestica L.) germplasm of Southern Italy. Sustainability 2019, 11, 4112. [Google Scholar] [CrossRef] [Green Version]
- Krigas, N.; Tsoktouridis, G.; Anestis, I.; Khabbach, A.; Libiad, M.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Lamchouri, F.; Tsiripidis, I.; Tsiafouli, M.A.; et al. Exploring the potential of neglected local endemic plants of three Mediterranean regions in the ornamental sector: Value chain feasibility and readiness timescale for their sustainable exploitation. Sustainability 2021, 13, 2539. [Google Scholar] [CrossRef]
- Bourgou, S.; Ben Haj Jilani, I.; Karous, O.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Greveniotis, V.; et al. Medicinal-cosmetic potential of the local endemic plants of Crete (Greece), Northern Morocco and Tunisia: Priorities for conservation and sustainable exploitation of neglected and underutilized phytogenetic resources. Biology 2021, 10, 1344. [Google Scholar] [CrossRef]
- Karapatzak, E.; Krigas, N.; Ganopoulos, I.; Papanastasi, K.; Kyrkas, D.; Yfanti, P.; Nikisianis, N.; Karydas, A.; Manthos, I.; Kosma, I.S.; et al. Documenting Greek indigenous germplasm of cornelian cherry (Cornus mas L.) for sustainable utilization: Molecular authentication, asexual propagation, and phytochemical evaluation. Plants 2022, 11, 1345. [Google Scholar] [CrossRef]
- Accogli, R.; Tomaselli, V.; Direnzo, P.; Perrino, E.V.; Albanese, G.; Urbano, M.; Laghetti, G. Edible halophytes and halo-tolerant species in Apulia region (Southeastern Italy): Biogeography, traditional food use and potential sustainable crops. Plants 2023, 12, 549. [Google Scholar] [CrossRef]
- Perrino, E.V.; Tomaselli, V.; Wagensommer, R.P.; Silletti, G.N.; Esposito, A.; Stinca, A. Ophioglossum lusitanicum L.: New records of plant community and 92/43/EEC Habitat in Italy. Agronomy 2022, 12, 3188. [Google Scholar] [CrossRef]
- Cosmulescu, S.; Trandafir, I.; Nour, V. Phenolic acids and flavonoids profiles of extracts from edible wild fruits and their antioxidant properties. Int. J. Food Prop. 2017, 20, 3124–3134. [Google Scholar] [CrossRef] [Green Version]
- Che, C.-T.; Zhang, H. Plant Natural Products for Human Health. Int. J. Molec. Sci. 2019, 20, 830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloupa, E.; Karapatzak, E.; Ganopoulos, I.; Karydas, A.; Papanastasi, K.; Kyrkas, D.; Yfanti, P.; Nikisianis, N.; Zahariadis, A.; Kosma, I.S.; et al. Molecular authentication, phytochemical evaluation and asexual propagation of wild-growing Rosa canina L. (Rosaceae) genotypes of Northern Greece for sustainable exploitation. Plants 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Karapatzak, E.; Dichala, O.; Ganopoulos, I.; Karydas, A.; Papanastasi, K.; Kyrkas, D.; Yfanti, P.; Nikisianis, N.; Fotakis, D.; Patakioutas, G.; et al. Molecular authentication, propagation trials and field establishment of Greek native genotypes of Sambucus nigra L. (Caprifoliaceae): Setting the basis for domestication and sustainable utilization. Agronomy 2022, 12, 114. [Google Scholar] [CrossRef]
- Kostas, S.; Hatzilazarou, S.; Pipinis, E.; Bourgou, S.; Ben Haj Jilani, I.; Ben Othman, W.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; et al. DNA barcoding, GIS-facilitated seed germination and pilot cultivation of Teucrium luteum subsp. gabesianum (Lamiaceae), a Tunisian local endemic with potential medicinal and ornamental value. Biology 2022, 11, 462. [Google Scholar] [CrossRef]
- Kazankaya, A.; Yörük, E.; Doğan, A. Effect of IBA on rooting of Rosa canina hardwood cuttings from lake Van region, Turkey. Acta Hort. 2005, 690, 153–158. [Google Scholar] [CrossRef]
- Hassanpour, H.; Ali Shiri, M. Propagation of Iranian cornelian cherry (Cornus mas L.) by rooted stem cuttings. Not. Sci. Biol. 2014, 6, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Pradeep Kumar, K.; Prabhat Kumar, D.; Pradipta Kumar, M.; Pratap Chandra, P. Effect of auxins on rooting of stem cuttings in Hypericum gaitii. J. Herbs Spices Medic. Plants 2020, 26, 423–434. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Krigas, N.; Sarropoulou, V.; Maloupa, E.; Tsoktouridis, G. Propagation and ex-situ conservation of Lomelosia minoana subsp. minoana and Scutellaria hirta—Two ornamental and medicinal Cretan endemics (Greece). Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12168. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; El Haissoufi, M.; Pipinis, E.; Kostas, S.; Libiad, M.; Khabbach, A.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; et al. GIS-Facilitated seed germination and multifaceted evaluation of the Endangered Abies marocana Trab. (Pinaceae) enabling conservation and sustainable exploitation. Plants 2021, 10, 2606. [Google Scholar] [CrossRef]
- Paschalidis, K.; Fanourakis, D.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Kalogiannakis, K.; Debouba, F.J.; Ipsilantis, I.; Tsoktouridis, G.; et al. Pilot Cultivation of the Vulnerable Cretan endemic Verbascum arcturus L. (Scrophulariaceae): Effect of fertilization on growth and quality features. Sustainability 2021, 13, 14030. [Google Scholar] [CrossRef]
- Fanourakis, D.; Paschalidis, K.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Liapaki, E.; Jouini, M.; Ipsilantis, I.; Maloupa, E.; et al. Pilot cultivation of the local endemic Cretan marjoram Origanum microphyllum (Benth.) Vogel (Lamiaceae): Effect of fertilizers on growth and herbal quality features. Agronomy 2022, 12, 94. [Google Scholar] [CrossRef]
- Pipinis, E.; Hatzilazarou, S.; Kostas, S.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; et al. Facilitating conservation and bridging gaps for the sustainable exploitation of the Tunisian local endemic plant Marrubium aschersonii (Lamiaceae). Sustainability 2022, 14, 1637. [Google Scholar] [CrossRef]
- Morugán-Coronado, A.; Linares, C.; Gómez-López, M.D.; Faz, Á.; Zornoza, R. The impact of intercropping, tillage and fertilizer type on soil and crop yield in fruit orchards under Mediterranean conditions: A meta-analysis of field studies. Agric. Syst. 2020, 178, 102736. [Google Scholar] [CrossRef]
- Hu, Y.; Zhan, P.; Thomas, B.W.; Zhao, J.; Zhang, X.; Yan, H.; Zhang, Z.; Chen, S.; Shi, X.; Zhang, Y. Organic carbon and nitrogen accumulation in orchard soil with organic fertilization and cover crop management: A global meta-analysis. Sci. Total Environ. 2022, 852, 158402. [Google Scholar] [CrossRef]
- Lombardo, E.; Bancheva, S.; Domina, G.; Venturella, G. Distribution, ecological role and symbioses of selected shrubby species in the Mediterranean Basin: A review. Plant Biosyst. 2020, 154, 438–454. [Google Scholar] [CrossRef]
- Boratynski, A.; Browicz, K.; Zieliński, J. Chorology of Trees and Shrubs in Greece; Polish Academy of Sciences: Poznan/Kornik, Poland, 1992. [Google Scholar]
- Ganeva, T.; Uzunova, K. Leaf epidermis structure in Amelanchier ovalis Medic. (Rosaceae). Biotechnol. Biotechnol. Equip. 2010, 24, 36–38. [Google Scholar] [CrossRef] [Green Version]
- Zadernowski, R.; Naczk, M.; Nesterowicz, J. Phenolic acid profiles in some small berries. J. Agric. Food Chem. 2005, 53, 2118–2124. [Google Scholar] [CrossRef]
- Ochmian, I.; Kubus, M.; Dobrowolska, A. Description of plants and assessment of chemical properties of three species from the Amelanchier genus. Dendrobiology 2013, 70, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Asyakina, L.; Atuchin, V.; Drozdova, M.; Kozlova, O.; Prosekov, A. Ex vivo and in vitro antiaging and antioxidant extract activity of the Amelanchier ovalis from Siberia. Int. J. Molec. Sci. 2022, 23, 15156. [Google Scholar] [CrossRef]
- Donno, D.; Mellano, M.G.; Cerutti, A.K.; Beccaro, G.L. Nutraceuticals in Alternative and Underutilized Fruits as Functional Food Ingredients: Ancient Species for New Health Needs. In Handbook of Food Bioengineering, 17, Alternative and Replacement Foods; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Jantová, S.; Nagy, M.; Ruzěková, L.; Grancăi, D. Antibacterial activity of plant extracts from the families Fabaceae, Oleaceae, Philadelphaceae, Rosaceae and Staphyleaceae. Phytother. Res. 2000, 14, 601–603. [Google Scholar] [CrossRef]
- Laksaeva, E.A. Fruits of plants of Amelanchier genus (Amelanchier Medic.) as source of biologically active substances and minerals. I. P. Pavlov Russ. Med. Biol. Herald 2018, 26, 296–304. [Google Scholar] [CrossRef]
- Zazharskyi, V.V.; Davydenko, P.O.; Kulishenko, O.M.; Borovik, I.V.; Brygadyrenko, V.V. Antimicrobial activity of 50 plant extracts. Biosyst. Divers. 2019, 27, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Bishop, B.H.; Nelson, S.H. Propagation and transplanting of Saskatoon (Amelanchier alnifolia Nutt.) softwood cuttings. Can. J. Plant Sci. 1980, 60, 883–890. [Google Scholar] [CrossRef]
- Still, S.M.; Zanon, S. Effects of K-IBA rates and timing on rooting percentage and root quality of Amelanchier laevis. J. Environ. Hortic. 1991, 9, 86–88. [Google Scholar] [CrossRef]
- Bounous, G.; Bullano, F.; Peano, C. Propagation by softwood cuttings of Amelanchier canadensis Medic., Cornus mas L., Elaeagnus umbellate Thunb. and Hippophae rhamnoides L. Turin University (Italy). Istituto di Coltivazioni Arboree. Monti Boschi 1992, 43, 51–57. [Google Scholar]
- Milla, R.; Castro-Díez, P.; Maestro-Martínez, M.; Montserrat-Martí, G. Environmental constraints on phenology and internal nutrient cycling in the Mediterranean winter-deciduous shrub Amelanchier ovalis Medicus. Plant Biol. 2005, 7, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Milla, R.; Castro-Díez, P.; Montserrat-Martí, G. Phenology of Mediterranean woody plants from NE Spain: Synchrony, seasonality, and relationships among phenophases. Flora 2010, 205, 190–199. [Google Scholar] [CrossRef]
- Da Costa, C.T.; de Almeida, M.R.; Ruedell, C.M.; Schwambach, J.; Maraschin, F.S.; Fett-Neto, A.G. When stress and development go hand in hand: Main hormonal controls of adventitious rooting in cuttings. Front. Plant Sci. 2013, 4, 133. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chen, Y.; Jiang, C.; Lu, M.-Z.; Zhang, J. Exogenous hormones supplementation improve adventitious root formation in woody plants. Front. Bioeng. Biotechnol. 2022, 10, 1009531. [Google Scholar] [CrossRef]
- Bryant, P.H.; Trueman, S.J. Stem anatomy and adventitious root formation in cuttings of Angophora, Corymbia and Eucalyptus. Forests 2015, 6, 1227–1238. [Google Scholar] [CrossRef]
- Pacholczak, A.; Jędrzejuk, A.; Sobczak, M. Shading and natural rooting biostimulator enhance potential for vegetative propagation of dogwood plants (Cornus alba L.) via stem cuttings. S. Afr. J. Bot. 2017, 109, 34–41. [Google Scholar] [CrossRef]
- Opeña, J.M.; Sotto, R.C.; Salazar, B.M.; Protacio, C.M. Morpho-anatomical investigation on the adventitious rooting of hard-to-root Excelsa coffee (Coffea excelsa A. Chev.) stem cuttings. Philip. Agric. Sci. 2020, 103, 303–310. [Google Scholar]
- Liu, G.; Zhao, J.; Liao, T.; Wang, Y.; Guo, L.; Yao, Y.; Cao, J. Histological dissection of cutting-inducible adventitious rooting in Platycladus orientalis reveals developmental endogenous hormonal homeostasis. Ind. Crops Prod. 2021, 170, 113817. [Google Scholar] [CrossRef]
- Balta, M.F.; Erol, I.U.; Özrenk, K.; Karakaya, O.; Uzun, S. Investigation on propagation with softwood cuttings of Cornelian cherry (Cornus mas L.) genotypes. Türkiye Tarımsal Araştırmalar Derg. 2019, 6, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.P.; Preece, J.E.; Aradhya, M.; Gradziel, T. Rooting response of Prunus wild relative semi-hardwood cuttings to indole-3-butyric acid potassium salt (KIBA). Sci. Hortic. 2020, 263, 109144. [Google Scholar] [CrossRef]
- Rosa, G.G.; da Zanandrea, I.; Mayer, N.A.; Bianchi, V.J. Effect of genotype on rooting and acclimatization of semi hardwood cutting of peach rootstock. Rev. Cienc. Agrovet. 2017, 16, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Uğur, R.; Gündeşli, M.A. Investigation of propagation with hardwood cutting in different Prunus species. Inter. J. Agric. For. Life Sci. 2020, 4, 184–189. Available online: https://dergipark.org.tr/en/pub/ijafls/issue/57325/782302 (accessed on 18 February 2023).
- Justamante, M.S.; Mhimdi, M.; Molina-Pérez, M.; Albacete, A.; Moreno, M.Á.; Mataix, I.; Pérez-Pérez, J.M. Effects of auxin (Indole-3-butyric Acid) on adventitious root formation in peach-based Prunus rootstocks. Plants 2022, 11, 913. [Google Scholar] [CrossRef]
- Nasri, F.; Fadakar, A.; Saba, M.K.; Yousefi, B. Study of indole butyric acid IBA effects on cutting rooting improving some of wild genotypes of damask roses Rosa damascena Mill. J. Agric. Sci. 2015, 60, 263–275. [Google Scholar] [CrossRef]
- Izadi, Z.; Zarei, H.; Alizadeh, M. Role of grafting technique on the success of stenting propagation of two rose (Rosa sp.) varieties. Am. J. Plant Sci. 2013, 4, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Bijelić, S.M.; Gološin, B.R.; Cerović, S.B.; Bogdanović, B.V. A comparison of grafting methods for the production of quality planting material of promising cornelian cherry selections (Cornus mas L.) in Serbia. J. Agric. Sci. Technol. 2016, 18, 223–231. Available online: http://jast.modares.ac.ir/article-23-1758-en.html (accessed on 18 February 2023).
- Cornescu, F.; Achim, G.; Cosmulescu, S. Vegetative propagation of Cornelian cherry (Cornus mas L.) selections. Not. Sci. Biol. 2020, 12, 836–841. [Google Scholar] [CrossRef]
- Druege, U.; Hilo, A.; Pérez-Pérez, J.M.; Klopotek, Y.; Acosta, M.; Shahinnia, F.; Zerche, S.; Franken, P.; Hajirezaei, M.R. Molecular and physiological control of adventitious rooting in cuttings: Phytohormone action meets resource allocation. Ann. Bot. 2019, 123, 929–949. [Google Scholar] [CrossRef] [Green Version]
- Ekin, H.N.; Gokbulut, A.; Aydin, Z.U.; Donmez, A.A.; Orhan, I.E. Insight into anticholinesterase and antioxidant potential of thirty-four Rosaceae samples and phenolic characterization of the active extracts by HPLC. Ind. Crops Prod. 2016, 91, 104–113. [Google Scholar] [CrossRef]
- Lavola, A.; Reijo Karjalainen, R.; Julkunen-Tiitto, R. Bioactive polyphenols in leaves, stems, and berries of Saskatoon (Amelanchier alnifolia Nutt.) cultivars. J. Agric. Food Chem. 2012, 60, 1020–1027. [Google Scholar] [CrossRef]
- Tian, Y.; Puganen, A.; Alakomi, H.-L.; Uusitupa, A.; Saarela, M.; Yang, B. Antioxidative and antibacterial activities of aqueous ethanol extracts of berries, leaves, and branches of berry plants. Food Res. Int. 2018, 106, 291–303. [Google Scholar] [CrossRef]
- Grygorieva, O.; Vergun, O.; Klymenko, S.; Zhurba, M.; Horčinová Sedláčková, V.; Ivanišová, E.; Brindza, J. Estimation of phenolic compounds content and antioxidant activity of leaves extracts of some selected non-traditional plants. Potravin. Slovak J. Food Sci. 2020, 14, 501–509. [Google Scholar] [CrossRef]
- Mȩczarska, K.; Cyboran-Mikołajczyk, S.; Włoch, A.; Bonarska-Kujawa, D.; Oszmiański, J.; Kleszczyńska, H. Polyphenol content and bioactivity of Saskatoon (Amelanchier alnifolia Nutt.) leaves and berries. Acta Pol. Pharm. Drug Res. 2017, 74, 660–669. Available online: https://www.ptfarm.pl/pub/File/Acta_Poloniae/2017/2/660.pdf (accessed on 31 January 2023).
- Zengin, G.; Diuzheva, A.; Jekő, J.; Cziáky, Z.; Bulut, G.; Dogan, A.; Haznedaroglu, M.Z.; Rengasamy, K.R.R.; Lobine, D.; Bahadori, M.B.; et al. HPLC–MS/MS-based metabolic profiling and pharmacological properties of extracts and infusion obtained from Amelanchier parviflora var. dentata. Ind. Crops Prod. 2018, 124, 699–706. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Eisner, P. How does the phenol structure influence the results of the Folin-Ciocalteu assay? Antioxidants 2021, 10, 811. [Google Scholar] [CrossRef] [PubMed]
- Krivoruchko, E.V.; Samoilova, V.A.; Demeshko, O.V. Carboxylic acids and essential oil from leaves of Amelanchier ovalis. Chem. Nat. Comp. 2021, 57, 578–579. [Google Scholar] [CrossRef]
- Green, R.C.; Mazza, G. Relationships between anthocyanins, total phenolics, carbohydrates, acidity and colour of Saskatoon berries. Can. Inst. Food Sci. Technol. J. 1986, 19, 107–113. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Knowles, N.R. Physical and chemical changes during growth, maturation, and ripening of saskatoon (Amelanchier alnifolia) fruit. Can. J. Bot. 1997, 75, 1215–1225. [Google Scholar] [CrossRef]
- Bernal, M.; Estiarte, M.; Peñuelas, J. Drought advances spring growth phenology of the Mediterranean shrub Erica multiflora. Plant Biol. 2011, 13, 252–257. [Google Scholar] [CrossRef]
- Rosati, A.; Paoletti, A.; Al Hariri, R.; Morelli, A.; Famiani, F. Resource investments in reproductive growth proportionately limit investments in whole-tree vegetative growth in young olive trees with varying crop loads. Tree Physiol. 2018, 38, 1267–1277. [Google Scholar] [CrossRef] [Green Version]
- Arrobas, M.; Ribeiro, A.; Barreales, D.; Pereira, E.L.; Rodrigues, M.Â. Soil and foliar nitrogen and boron fertilization of almond trees grown under rainfed conditions. Europ. J. Agron. 2019, 106, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Sete, P.B.; Comin, J.J.; Ciotta, M.N.; Salume, J.A.; Thewes, F.; Brackmann, A.; Toselli, M.; Nava, G.; Rozane, D.E.; Loss, A.; et al. Nitrogen fertilization affects yield and fruit quality in pear. Sci. Hortic. 2019, 258, 108782. [Google Scholar] [CrossRef]
- Martínez-Mena, M.; Garcia-Franco, N.; Almagro, M.; Ruiz-Navarro, A.; Albaladejo, J.; de Aguilar, J.M.; Gonzalez, D.; Querejeta, J.I. Decreased foliar nitrogen and crop yield in organic rainfed almond trees during transition from reduced tillage to no-tillage in a dryland farming system. Europ. J. Agron. 2013, 49, 149–157. [Google Scholar] [CrossRef]
- Chatzistathis, T.; Papadakis, I.E.; Papaioannou, A.; Chatzissavvidis, C.; Giannakoula, A. Comparative study effects between manure application and a controlled-release fertilizer on the growth, nutrient uptake, photosystem II activity and photosynthetic rate of Olea europaea L. (cv. ‘Koroneiki’). Sci. Hortic. 2020, 264, 109176. [Google Scholar] [CrossRef]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and vine water deficits as factors in wine grape composition: Anthocyanins and tannins. Aust. J. Grape Wine Res. 2004, 10, 100–107. [Google Scholar] [CrossRef]
- Remberg, S.F.; Sønsteby, A.; Aaby, K.; Heide, O.M. Influence of post flowering temperature on fruit size and chemical composition of Glen Ample raspberry (Rubus idaeus L.). J. Agric. Food Chem. 2010, 58, 9120–9128. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Mantri, N.; Lou, H.; Hu, Y.; Sun, D.; Zhu, Y.; Dong, T.; Lu, H. Effects of elevated CO2 and temperature on yield and fruit quality of Strawberry (Fragaria × ananassa Duch.) at two levels of nitrogen application. PLoS ONE 2012, 7, e41000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lwin, H.P.; Lee, J. Fruit quality and major metabolites in cold-stored ‘Wonhwang’ Asian pears are differentially affected by fruit size. J. Sci. Food Agric. 2020, 100, 5117–5125. [Google Scholar] [CrossRef] [PubMed]
- Strid, A. Atlas of the Aegean Flora, Part 1: Text & Plates; Part 2: Maps; Englera, Volume 33; Botanic Garden and Botanical Museum Berlin, Freie Universität Berlin: Berlin, Germany, 2016; ISBN1 978-3-921800-97-3. ISBN2 978-3-921800-98-0. [Google Scholar]
- Sidiropoulou, E.; Marugán-Hernández, V.; Skoufos, I.; Giannenas, I.; Bonos, E.; Aguiar-Martins, K.; Lazari, D.; Papagrigoriou, T.; Fotou, K.; Grigoriadou, K.; et al. In vitro antioxidant, antimicrobial, anticoccidial, and anti-inflammatory study of essential oils of oregano, thyme, and sage from Epirus, Greece. Life 2022, 12, 1783. [Google Scholar] [CrossRef] [PubMed]
- Risaliti, L.; Kehagia, A.; Daoultzi, E.; Lazari, D.; Bergonzi, M.C.; Vergkizi-Nikolakaki, S.; Hadjipavlou-Litina, D.; Bilia, A.R. Liposomes loaded with Salvia triloba and Rosmarinus officinalis essential oils: In vitro assessment of antioxidant, anti-inflammatory and antibacterial activities. J. Drug Deliv. Sci. Technol. 2019, 51, 493–498. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Hormone Treatment | (%) Rooting | Mean Root Number * | Mean Root Length (mm) |
|---|---|---|---|---|
| 1 | Control | 0.0 | - | - |
| 2000 ppm IBA | 66.6 | 6.25 (±0.75) a | 31.27 (±5.88) A | |
| 4000 ppm IBA | 83.3 † | 6.40 (±2.03) a | 22.26 (±4.23) A | |
| 6000 ppm IBA | 50.0 | 6.66 (±0.33) a | 34.74 (±6.13) A | |
| 2500 ppm NAA | 83.3 † | 6.20 (±0.58) a | 24.24 (±3.27) A | |
| 5000 ppm NAA | 66.6 | 36.50 (±16.65) b | 18.42 (±3.69) A | |
| 2 | Control | 0.0 | - | - |
| 2000 ppm IBA | 16.6 | 6.00 (±0.00) ** | 33.50 (±0.00) | |
| 4000 ppm IBA | 33.3 † | 11.00 (±0.00) | 31.18 (±4.54) A | |
| 6000 ppm IBA | 0.0 | - | - | |
| 2500 ppm NAA | 33.3 † | 9.00 (±0.00) | 18.11 (±2.66) A | |
| 5000 ppm NAA | 0.0 | - | - |
| Plant Organ | Phytochemical Attribute | |
|---|---|---|
| Total Phenolic Content (mg GAE L−1) | Antioxidant Activity (%RSA) | |
| Leaves | 37.639 (4.432) a | 93.239 (1.609) a |
| Twigs | 32.476 (3.966) a | 93.431 (0.626) a |
| Flowers | 9.468 (1.675) b | 92.056 (1.596) a |
| Young fruits | 33.214 (3.108) a | 93.136 (1.541) a |
| No | IPEN Accession Number | Greek Prefecture | Area | Coordinates (HGRS87/EGSA87) (Lat, Lon) | Altitude (m) | Collected Material * |
|---|---|---|---|---|---|---|
| 1 | GR-1-BBGK-19,185 | Thessaly | Mt Lakmos, Trikala | 39.667963, 21.135741 | 1984 | HC |
| 2 | GR-1-BBGK-19,612 | Thessaly | Mt Lakmos, Trikala | 39.650822, 21.150658 | 1763 | SC |
| 3 | GR-1-BBGK-19,613 | Thessaly | Venetikos river, Trikala | 40.052361, 21.481166 | 468 | HC, SC |
| 4 | GR-1-BBGK-19,680 | Macedonia | Mt Paiko, Kilkis | 40.98824, 22.34423 | 750 | HC, SC |
| 5 | GR-1-BBGK-19,845 | Macedonia | Lake Prespa, Oxia | 40.73076, 21.12622 | 1196 | HC, SC |
| 6 | GR-1-BBGK-19,846 | Macedonia | Lake Prespa, Oxia | 40.72997, 21.11763 | 1249 | HC, SC |
| 7 | GR-1-BBGK-19,928 | Macedonia | Mt Tzena, Pella | 41.12088, 22.21743 | 1152 | HC, SC |
| 8 | GR-1-BBGK-19,929 | Macedonia | Mt Tzena, Pella | 41.12242, 22.21924 | 998 | HC, SC |
| 9 | GR-1-BBGK-19,987 | Macedonia | Lake Prespa, Oxia | 40.72767, 21.11599 | 1282 | HC, SC |
| 10 | GR-1-BBGK-04,2547 | Macedonia | Tzena, Pella ** | 41.123857, 22.184903 | 1120 | HC, SC |
| No | Pre-Treatment | Treatment |
|---|---|---|
| 1 | - | Control |
| 2 | - | 2000 ppm IBA |
| 3 | - | 4000 ppm IBA |
| 4 | - | 6000 ppm IBA |
| 5 | - | 2500 ppm NAA |
| 6 | - | 5000 ppm NAA |
| 7 | 5% OFS * | 2000 ppm IBA |
| 8 | 5% OFS * | 4000 ppm IBA |
| 9 | 5% OFS * | 2500 ppm NAA |
| 10 | 5% OFS * | 5000 ppm NAA |
| 11 | 10% OFS * | 2000 ppm IBA |
| 12 | 10% OFS * | 4000 ppm IBA |
| 13 | - | 0.25% powder ΙΒA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karapatzak, E.; Papagrigoriou, T.; Papanastasi, K.; Dichala, O.; Karydas, A.; Nikisianis, N.; Patakioutas, G.; Lazari, D.; Krigas, N.; Maloupa, E. From the Wild to the Field: Documentation, Propagation, Pilot Cultivation, Fertilization, and Phytochemical Evaluation of the Neglected and Underutilized Amelanchier ovalis Medik. (Rosaceae). Plants 2023, 12, 1142. https://doi.org/10.3390/plants12051142
Karapatzak E, Papagrigoriou T, Papanastasi K, Dichala O, Karydas A, Nikisianis N, Patakioutas G, Lazari D, Krigas N, Maloupa E. From the Wild to the Field: Documentation, Propagation, Pilot Cultivation, Fertilization, and Phytochemical Evaluation of the Neglected and Underutilized Amelanchier ovalis Medik. (Rosaceae). Plants. 2023; 12(5):1142. https://doi.org/10.3390/plants12051142
Chicago/Turabian StyleKarapatzak, Eleftherios, Theodora Papagrigoriou, Katerina Papanastasi, Olga Dichala, Antonis Karydas, Nikos Nikisianis, Giorgos Patakioutas, Diamanto Lazari, Nikos Krigas, and Eleni Maloupa. 2023. "From the Wild to the Field: Documentation, Propagation, Pilot Cultivation, Fertilization, and Phytochemical Evaluation of the Neglected and Underutilized Amelanchier ovalis Medik. (Rosaceae)" Plants 12, no. 5: 1142. https://doi.org/10.3390/plants12051142