

In Vitro Fecal Fermentation of Euphorbia humifusa-Derived Polysaccharides and Their Protective Effect against Ulcerative Colitis in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction and Purification of EHPs
2.3. Structural Characterization of EHPs
2.3.1. Chemical Composition
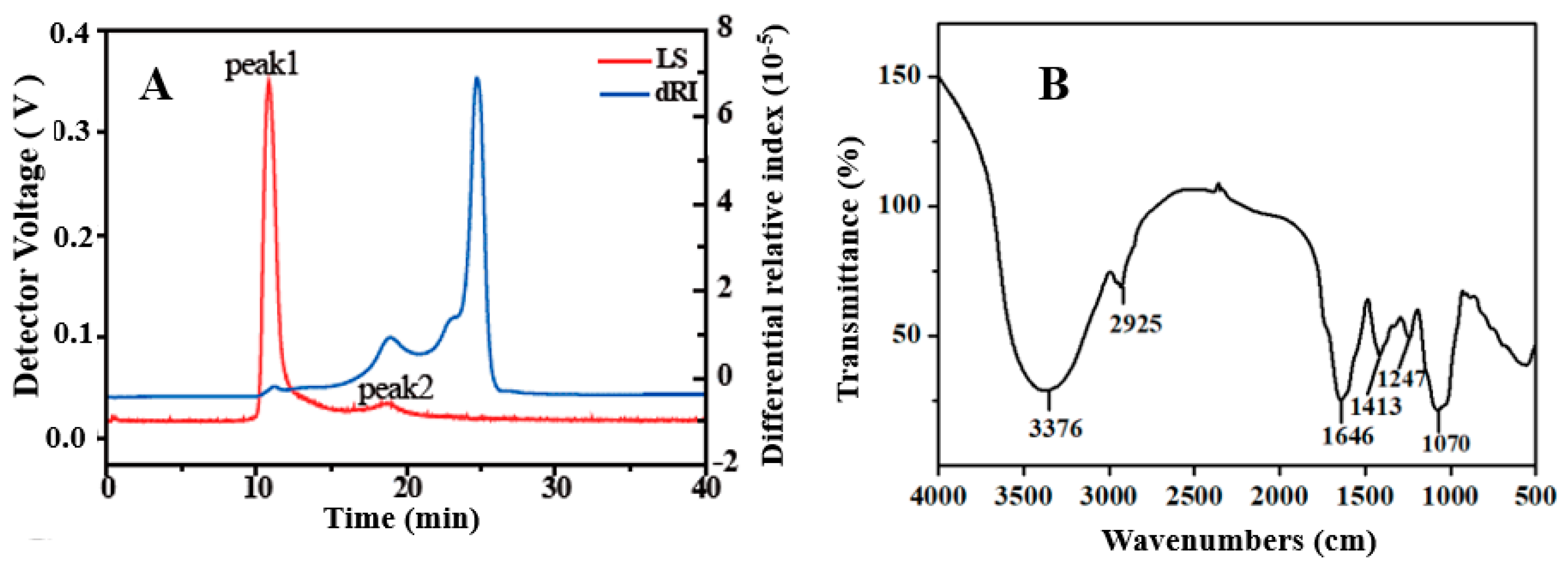
2.3.2. Molecular Weight (Mw) Distribution
2.3.3. Monosaccharide Composition
2.3.4. Fourier Transform Infrared (FTIR) Spectroscopic Analysis
2.4. In Vitro Absorption of EHPs
2.4.1. Fluorescent Labeling of Polysaccharides
2.4.2. Caco-2 Cell-Based Intestinal Absorption Model
2.4.3. EHP Uptake by Caco-2 Cells
2.5. In Vitro Fermentation of EHPs
2.5.1. Fermentation
2.5.2. Determination of SCFA Concentration
2.5.3. DNA Extraction and Sequence Analysis
2.6. Animal Studies
2.6.1. Experimental Design
2.6.2. qRT-PCR
2.7. Statistical Data Analysis
3. Results
3.1. Structural Characterization of EHPs
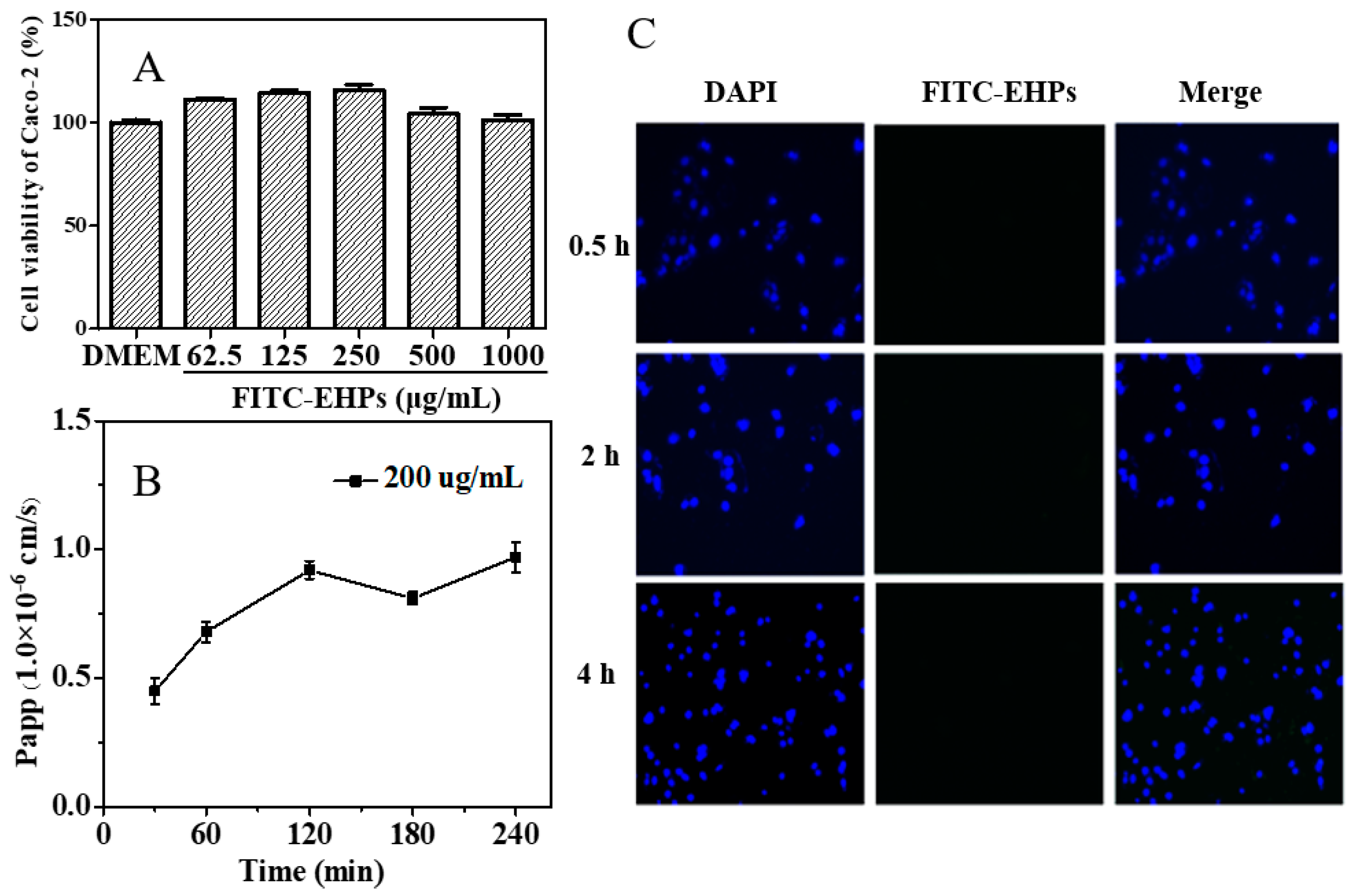
3.2. In Vitro Absorption Characteristics of Polysaccharides
3.3. In Vitro Fermentation of EHPs
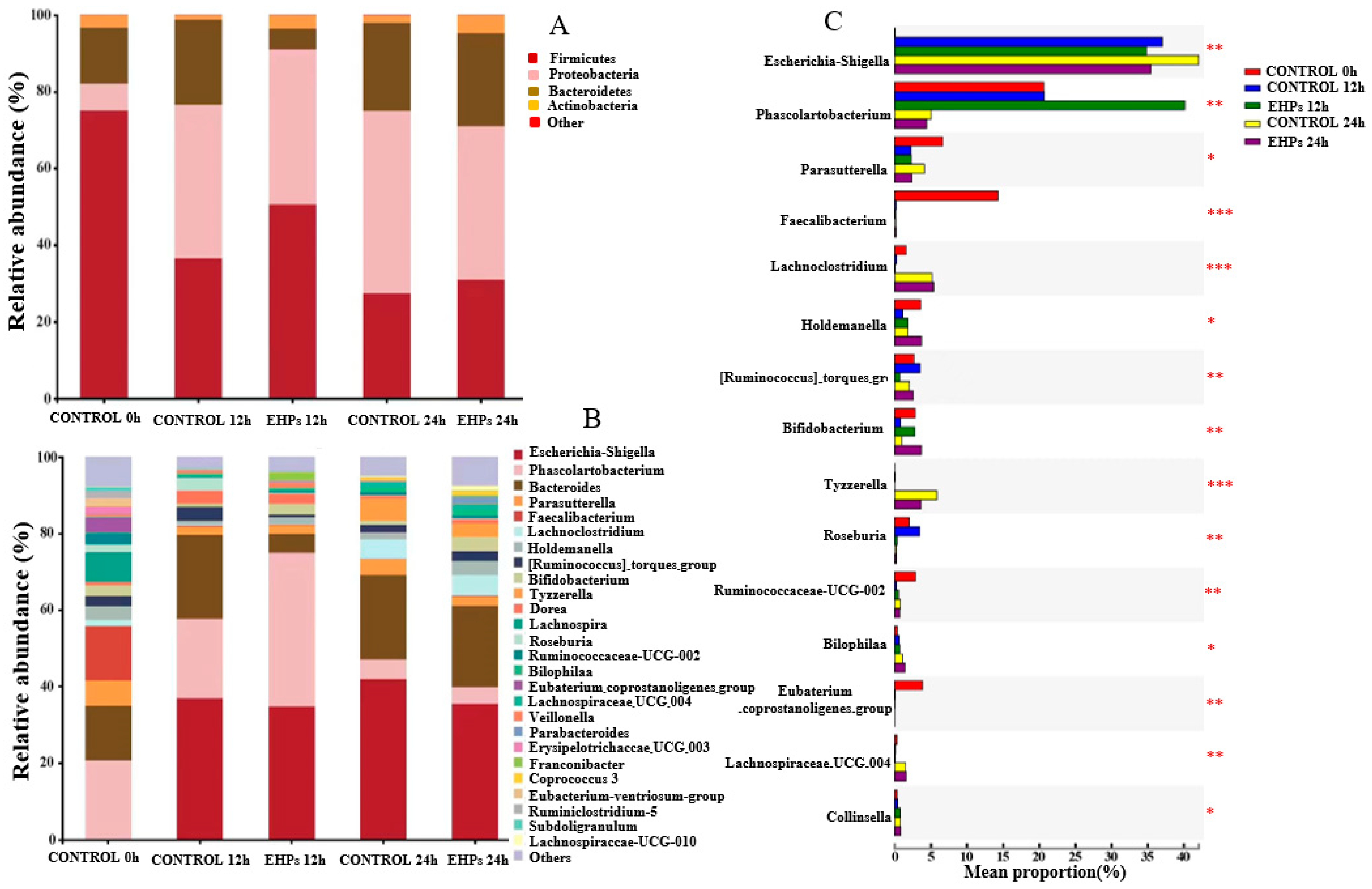
3.3.1. Changes in Intestinal Microecology
3.3.2. Changes in SCFA Content
3.4. Therapeutic Effects of EHP Treatment in DSS-Induced Colitis Mice
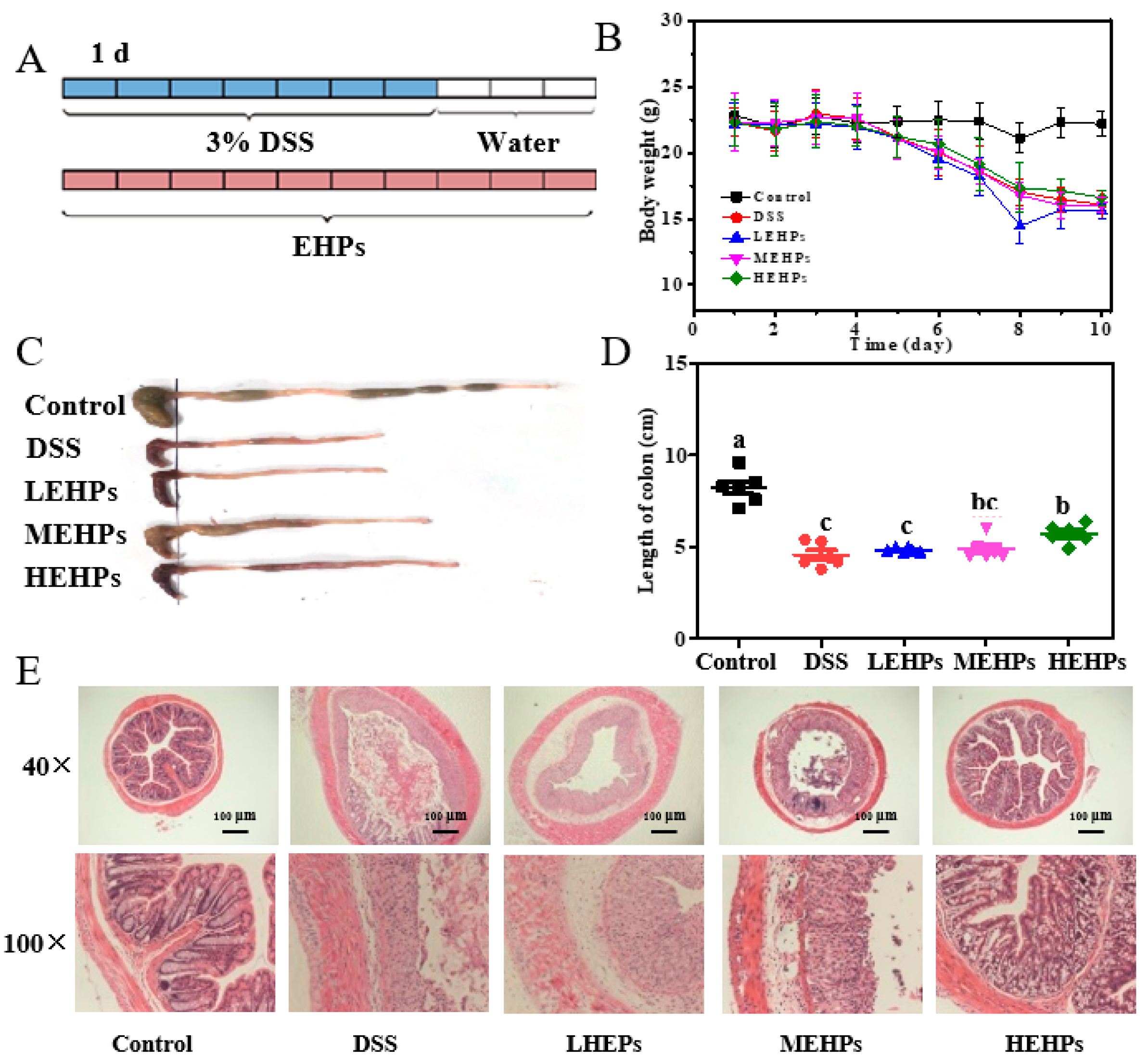
3.4.1. EHP Supplementation Ameliorated Colitis Symptoms
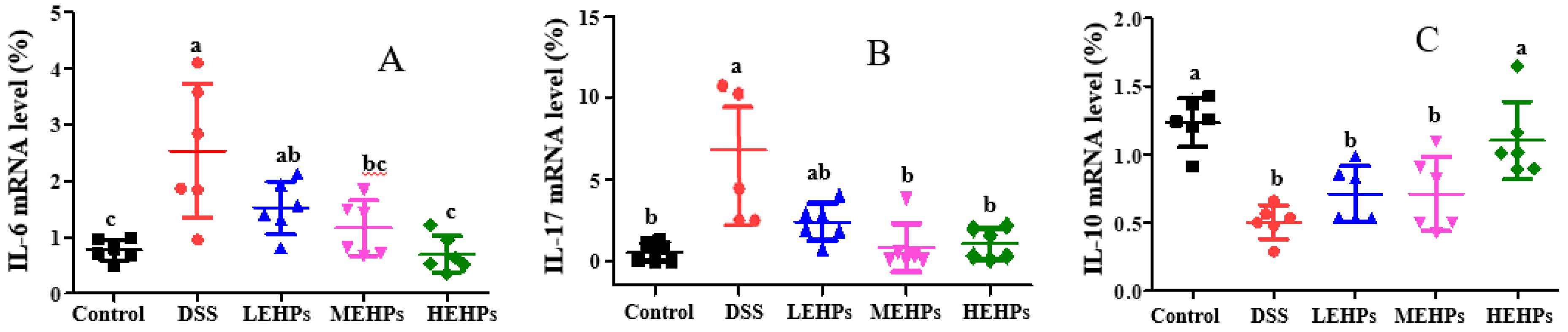
3.4.2. EHP Treatment Inhibited Colonic Inflammation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Papp | Apparent permeability coefficient |
| Ara | Arabinose |
| DSS | Dextran sulfate sodium |
| EHPs | Euphorbia humifusa-derived polysaccharides |
| FTIR | Fourier transform infrared spectroscopy |
| FITC | Fluorescein isothiocyanate |
| HPSEC | High-performance size exclusion chromatography |
| H&E | Hematoxylin-eosin |
| Gal | Galactose |
| Glu | Glucose |
| GluA | Glucuronic acid |
| Man | Mannose |
| SCFAs | Short-chain fatty acids |
| TEER | Transepithelial electrical resistance |
| UC | Ulcerative colitis |
References
- Chang, J.T. Pathophysiology of inflammatory bowel diseases. N. Engl. J Med. 2020, 383, 2652–2664. [Google Scholar] [CrossRef] [PubMed]
- Kayal, M.; Shah, S. Ulcerative colitis: Current and emerging treatment strategies. J. Clin. Med. 2019, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, A.; Miao, J.; Sun, H.; Yan, G.; Wu, F.; Wang, X. Gut microbiota as important modulator of metabolism in health and disease. RSC Adv. 2018, 8, 42380–42389. [Google Scholar] [CrossRef]
- Fei, Y.; Chen, Z.; Han, S.; Zhang, S.; Zhang, T.; Lu, Y.; Berglund, B.; Xiao, H.; Li, L.; Yao, M. Role of prebiotics in enhancing the function of next-generation probiotics in gut microbiota. Crit. Rev. Food Sci. Nutr. 2021. [Google Scholar] [CrossRef]
- Patnode, M.L.; Beller, Z.W.; Han, N.D.; Cheng, J.; Peters, S.L.; Terrapon, N.; Henrissat, B.; Gall, S.L.; Saulnier, L.; Hayashi, D.K.; et al. Interspecies competition impacts targeted manipulation of human gut bacteria by fiber-derived glycans. Cell 2019, 179, 59–73. [Google Scholar] [CrossRef]
- Han, R.; Pang, D.; Wen, L.; You, L.; Huang, R.; Kulikouskaya, V. In vitro digestibility and prebiotic activities of a sulfated polysaccharide from Gracilaria Lemaneiformis. J. Funct. Foods. 2020, 64, 103652. [Google Scholar] [CrossRef]
- Wu, D.T.; Nie, X.R.; Gan, R.Y.; Guo, H.; Fu, Y.; Yuan, Q.; Zhang, Q.; Qin, W. In vitro digestion and fecal fermentation behaviors of a pectic polysaccharide from okra (Abelmoschus esculentus) and its impacts on human gut microbiota. Food Hydrocolloids. 2021, 114, 106577. [Google Scholar] [CrossRef]
- Yuan, D.; Li, C.; Huang, Q.; Fu, X.; Dong, H. Current advances in the anti-inflammatory effects and mechanisms of natural polysaccharides. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- Shin, S.Y.; Kim, C.G.; Jung, Y.J.; Jung, Y.; Jung, H.; Im, J.; Lim, Y.; Lee, Y.H. Euphorbia humifusa Willd exerts inhibition of breast cancer cell invasion and metastasis through inhibition of TNFα-induced MMP-9 expression. BMC Complem. Altern. Med. 2016, 16, 413. [Google Scholar] [CrossRef] [Green Version]
- Rakotondrabe, T.F.; Fan, M.; Guo, M. Exploring potential antidiabetic and anti-inflammatory flavonoids from Euphorbia humifusa with an integrated strategy. Front. Pharmacol. 2022, 13, 980945. [Google Scholar] [CrossRef]
- Ding, Q.; Nie, S.; Hu, J.; Zong, X.; Li, Q.; Xie, M. In vitro and in vivo gastrointestinal digestion and fermentation of the polysaccharide from Ganoderma atrum. Food Hydrocolloids 2017, 63, 646–655. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.; Hamilton, J.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Chen, F.; Huang, G.; Yang, Z.; Hou, Y. Antioxidant activity of Momordica charantia polysaccharide and its derivatives. Int. J. Biol. Macromol. 2019, 10, 673–680. [Google Scholar] [CrossRef]
- Xia, Y.G.; Yu, L.S.; Liang, J.; Yang, B.Y.; Kuang, H.X. Chromatography and mass spectrometry-based approaches for perception of polysaccharides in wild and cultured fruit bodies of Auricularia auricular-judae. Int. J. Biol. Macromol. 2019, 137, 1232–1244. [Google Scholar] [CrossRef]
- Hu, Y.C.; Hu, J.L.; Li, J.; Wang, J.; Zhang, X.Y.; Wu, X.Y.; Li, X.; Guo, Z.N.; Zou, L.; Wu, D.T. Physicochemical characteristics and biological activities of soluble dietary fibers isolated from the leaves of different quinoa cultivars. Food. Res. Int. 2023, 163, 112166. [Google Scholar] [CrossRef]
- Wu, D.T.; An, L.Y.; Liu, W.; Hu, Y.C.; Wang, S.P.; Zou, L. In vitro fecal fermentation properties of polysaccharides from Tremella fuciformis and related modulation effects on gut microbiota. Food. Res. Int. 2022, 156, 111185. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Shen, Y.; Zhao, X.; Wang, X.; Wang, J.; Fan, K.; Zhan, X. Characterization of a novel polysaccharide from Ganoderma lucidum and its absorption mechanism in caco-2 cells and mice model. Int. J. Biol. Macromol. 2018, 118, 320–326. [Google Scholar] [CrossRef]
- Chen, S.; Chen, Z.; Wang, Y.; Hao, W.; Yuan, Q.; Zhou, H.; Gao, C.; Wang, Y.; Wu, X.; Wang, S. Targeted delivery of Chinese herb pair-based berberine/tannin acid self-assemblies for the treatment of ulcerative colitis. J. Adv. Res. 2022, 40, 263–267. [Google Scholar] [CrossRef]
- Ren, B.; Yuan, T.; Zhang, X.; Wang, L.; Pan, J.; Liu, Y.; Zhao, B.; Zhao, W.; Liu, Z.; Liu, X. Protective effects of sesamol on systemic inflammation and cognitive impairment in aging mice. J. Agric. Food. Chem. 2020, 68, 3099–3111. [Google Scholar] [CrossRef]
- Wang, J.; Geng, T.; Zou, Q.; Yang, N.; Zhao, W.; Li, Y.; Tan, X.; Yuan, T.; Xuebo, L.; Liu, Z. Lycopene prevents lipid accumulation in hepatocytes by stimulating PPARα and improving mitochondrial function. J. Funct. Foods 2020, 67, 103857. [Google Scholar] [CrossRef]
- Gu, J.Y.; Zhang, H.; Yao, H.; Zhou, J.; Duan, Y.Q.; Ma, H.L. Comparison of characterization, antioxidant and immunological activities of three polysaccharides from Sagittaria sagittifolia L. Carbohydr. Polym. 2020, 235, 115939. [Google Scholar] [CrossRef] [PubMed]
- Olawuyi, I.F.; Lee, W.Y. Structural characterization, functional properties and antioxidant activities of polysaccharide extract obtained from okra leaves (Abelmoschus esculentus). Food Chem. 2021, 354, 129437. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Wang, H.; Xia, M.; Deng, B.; Shen, H.; Ji, G.; Li, G.; Xie, Y. The effect of phytic acid on tight junctions in the human intestinal Caco-2 cell line and its mechanism. Eur. J. Pharm. Sci. 2015, 80, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.F.; Zhang, W.J.; Li, Q.; Zhao, J.; Feng, W.; Zhao, T.; Mao, G.; Chen, Y.; Xu, X.; Yang, L.; et al. Investigation of the uptake and transport of polysaccharide from Se-enriched Grifola frondosa in Caco-2 cells model. Int. J. Biol. Macromol. 2020, 158, 1330–1341. [Google Scholar] [CrossRef]
- Liu, C.; Kou, Y.; Zhang, X.; Cheng, H.; Chen, X.; Mao, S. Strategies and industrial perspectives to improve oral absorption of biological macromolecules. Expert Opin. Drug Deliv. 2018, 15, 223–233. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Dou, P.; Wu, Z.; Zheng, Z.; Pan, X.; Zhou, T.; Wang, K.P. Oral absorption characteristics and mechanisms of a pectin-type polysaccharide from Smilax China L. across the intestinal epithelium. Carbohydr. Polym. 2021, 270, 118383. [Google Scholar] [CrossRef]
- Zheng, Z.; Pan, X.; Luo, L.; Zhang, Q.; Huang, X.; Liu, Y.; Wang, K.; Zhang, Y. Advances in oral absorption of polysaccharides: Mechanism, affecting factors, and improvement strategies. Carbohydr. Polym. 2022, 282, 119110. [Google Scholar] [CrossRef]
- Wang, K.; Cheng, F.; Pan, X.; Zhou, T.; Liu, X.; Zheng, Z.; Luo, L.; Zhang, Y. Investigation of the transport and absorption of Angelica sinensis polysaccharide through gastrointestinal tract both in vitro and in vivo. Drug Deliv. 2017, 24, 1360–1371. [Google Scholar] [CrossRef]
- Wu, D.T.; He, Y.; Yuan, Q.; Wang, S.P.; Gan, R.Y.; Hu, Y.C.; Zou, L. Effects of molecular weight and degree of branching on microbial fermentation characteristics of okra pectic-polysaccharide and its selective impact on gut microbial composition. Food Hydrocolloids 2022, 132, 107897. [Google Scholar] [CrossRef]
- Wang, M.; Wichienchot, S.; He, X.; Fu, X.; Huang, Q.; Zhang, B. In vitro colonic fermentation of dietary fibers: Fermentation rate, short-chain fatty acid production and changes in microbiota. Trends Food Sci. Tech. 2019, 88, 1–9. [Google Scholar] [CrossRef]
- Annunziata, G.; Arnone, A.; Ciampaglia, R.; Tenore, G.C.; Novellino, E. Fermentation of foods and beverages as a tool for increasing availability of bioactive compounds. Focus on short-chain fatty acids. Foods 2020, 8, 999. [Google Scholar] [CrossRef]
- Do, M.H.; Seo, Y.S.; Park, H.Y. Polysaccharides: Bowel health and gut microbiota. Crit. Rev. Food Sci. Nutr. 2020, 61, 1212–1224. [Google Scholar] [CrossRef]
- Wang, L.; Cen, S.; Wang, G.; Lee, Y.K.; Zhao, J.; Zhang, H.; Chen, W. Acetic acid and butyric acid released in large intestine play different roles in the alleviation of constipation. J. Funct. Foods 2020, 69, 103953. [Google Scholar] [CrossRef]
- Qing, Y.; Xie, H.; Su, C.; Wang, Y.; Yu, Q.; Pang, Q.; Cui, F. Gut microbiome, short-chain fatty acids, and mucosa injury in young adults with human immunodeficiency virus infection. Dig. Dis. Sci. 2019, 64, 1830–1843. [Google Scholar] [CrossRef]
- Liu, Y.; Duan, X.; Duan, S.; Li, C.; Hu, B.; Liu, A.; Wu, Y.; Wu, H.; Chen, H.; Wu, W. Effects of in vitro digestion and fecal fermentation on stability and metabolic behavior of polysaccharides from Craterellus cornucopioides. Food Funct. 2020, 11, 6899–6910. [Google Scholar] [CrossRef]
- Hao, W.; Chen, Z.J.; Yuan, Q.; Ma, M.L.; Gao, C.F.; Zhou, Y.Y.; Zhou, H.F.; Wu, X.; Wu, D.T.; Farag, M.A. Ginger polysaccharides relieve ulcerative colitis via maintaining intestinal barrier integrity and gut microbiota modulation. Int. J. Biol. Macromol. 2022, 219, 730–739. [Google Scholar] [CrossRef]
- Nitima, T.; Waranya, C.; Kanokwan, J. Immune response and inflammatory pathway of ulcerative colitis. J. Basic Clin. Physiol. Pharmacol. 2019, 30, 1–10. [Google Scholar]
- Imaoka, A.; Shima, T.; Kato, K.; Mizuno, S.; Uehara, T.; Matsumoto, S.; Setoyama, H.; Hara, T.; Umesaki, Y. Anti-inflammatory activity of probiotic Bifidobacterium: Enhancement of IL-10 production in peripheral blood mononuclear cells from ulcerative colitis patients and inhibition of IL-8 secretion in HT-29 cells. World J. Gastroenterol. 2008, 14, 2511–2516. [Google Scholar] [CrossRef]
- Pan, X.; Yin, M.Y.; Guo, M.Z.; Niu, X.Y.; Han, L.R. The latest progress of natural food polysaccharides preventing ulcerative colitis by regulating intestinal microbiota. J. Funct. Foods 2022, 96, 105201. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, H.; Wang, X.; Yu, Y.; Xie, J. Natural food polysaccharides ameliorate inflammatory bowel disease and its mechanisms. Foods 2021, 10, 1288. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.J.; Wang, M.J.; Zeng, Z.Q.; Xie, M.H.; Xu, W.Q.; Peng, Y.J.; Zhou, W.T.; Sun, Y.; Zeng, X.X.; Liu, Z.H. Fuzhuan brick tea polysaccharides serve as a promising candidate for remodeling the gut microbiota from colitis subjects in vitro: Fermentation characteristic and anti-inflammatory activity. Food Chem. 2022, 391, 133203. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer | Reverse Primer |
|---|---|---|
| IL-6 | 5′-AGCGATGATGCACTGTCAGA-3′ | 5′-GGAACTCCAGAAGACCAGAGC-3′ |
| IL-17 | 5′-TTCTTTCAAACAAAGGACCAGC-3′ | 5′-GCAACCCAAGTAACCCTTAAAG-3′ |
| IL-10 | 5′-GACTTCACCATGGAACCCGT-3′ | 5′-GGAGACTGCCCATTCTCGAC-3′ |
| Items | Content |
|---|---|
| Composition (w%) | |
| Carbohydrate | 75.30 ± 0.04 |
| Protein | 2.02 ± 0.41 |
| Monosaccharide composition (molar ratio %) | |
| Mannose (Man) | 6.12 |
| Rhamnose (Rha) | 1.47 |
| Glucuronic acid (GluA) | 8.30 |
| Galacturonic acid (GalA) | 0.77 |
| Glucose (Glu) | 21.71 |
| Galactose (Gal) | 55.34 |
| Arabinose (Ara) | 6.30 |
| Sample | Time (h) | SCFAs(mmol/L) | ||||||
|---|---|---|---|---|---|---|---|---|
| Acetic Acid | Propionic Acid | Butyric Acid | Isobutyric Acid | Valeric Acid | Isovaleric Acid | Total | ||
| Control | 0 | ND | ND | ND | ND | ND | ND | ND |
| 4 | 1.21 ± 0.21 f | 1.51 ± 0.17 d | 0.18 ± 0.01 e | 0.07 ± 0.00 c | 0.07 ± 0.00 d | 0.05 ± 0.00 b | 3.09 ± 0.11 f | |
| 8 | 2.61 ± 0.09 e | 2.43 ± 0.29 cd | 1.53 ± 0.16 cd | 0.10 ± 0.01 c | 0.11 ± 0.01 cd | 0.05 ± 0.00 b | 6.63 ± 0.10 de | |
| 12 | 5.62 ± 0.26 c | 6.20 ± 0.35 b | 5.80 ± 0.67 b | 1.70 ± 0.18 b | 0.25 ± 0.01 bc | 0.16 ± 0.01 b | 19.73 ± 1.84 c | |
| 24 | 6.10 ± 0.46 c | 7.01 ± 0.30 b | 7.34 ± 0.27 a | 3.46 ± 0.39 a | 0.41 ± 0.02 b | 2.14 ± 0.12 a | 26.46 ± 2.55 b | |
| EHPs | 0 | ND | ND | ND | ND | ND | ND | ND |
| 4 | 1.64 ± 0.08 ef | 1.85 ± 0.09 d | 0.24 ± 0.01 de | 0.07 ± 0.01 c | 0.06 ± 0.00 d | 0.04 ± 0.00 b | 3.90 ± 0.67 ef | |
| 8 | 3.70 ± 0.42 d | 3.48 ± 0.35 c | 2.12 ± 0.42 c | 0.13 ± 0.01 c | 0.14 ± 0.01 cd | 0.07 ± 0.00 b | 9.64 ± 0.35 d | |
| 12 | 7. 50 ± 0.50 b | 7.20 ± 0.24 b | 5.43 ± 0.46 b | 1.81 ± 0.17 b | 0.35 ± 0.01 bc | 0.22 ± 0.02 b | 22.51 ± 1.81 c | |
| 24 | 8.82 ± 0.32 a | 9.92 ± 0.98 a | 7.62 ± 0.39 a | 3.24 ± 0.40 a | 0.90 ± 0.05 a | 2.25 ± 0.19 a | 32.75 ± 1.10 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, N.; Zhao, J.; Chang, S.; Li, S.; Liu, S.; Wang, C. In Vitro Fecal Fermentation of Euphorbia humifusa-Derived Polysaccharides and Their Protective Effect against Ulcerative Colitis in Mice. Foods 2023, 12, 751. https://doi.org/10.3390/foods12040751
Xiang N, Zhao J, Chang S, Li S, Liu S, Wang C. In Vitro Fecal Fermentation of Euphorbia humifusa-Derived Polysaccharides and Their Protective Effect against Ulcerative Colitis in Mice. Foods. 2023; 12(4):751. https://doi.org/10.3390/foods12040751
Chicago/Turabian StyleXiang, Ning, Jianbo Zhao, Siqiao Chang, Shasha Li, Shuwen Liu, and Chan Wang. 2023. "In Vitro Fecal Fermentation of Euphorbia humifusa-Derived Polysaccharides and Their Protective Effect against Ulcerative Colitis in Mice" Foods 12, no. 4: 751. https://doi.org/10.3390/foods12040751